
Генетическая несовместимость: Несовместимость партнеров при зачатии ребенка — что это такое, виды, признаки и что делать при несовместимости
Беременность при генетической несовместимости мужа и жены
Если супружеская пара очень долго не может зачать ребенка самостоятельно в течение 1 года необходимо обращение к специалистам. И кроме посещения врача-репродуктолога и уролога-андролога рекомендуется записаться и к врачу-генетику. Так как возможной причиной ненаступления беременности может являться генетическая несовместимость партнеров.
Такое сходство с точки зрения биологии и рождения детей – негативный фактор.
В случае генетической несовместимости даже при удачном оплодотворении возможно прерывание беременности или рождение ребенка с тяжелой патологией.
Тем не менее, возможности современной медицины позволяют найти альтернативные пути решения этой проблемы.
Что такое генетическая несовместимость и какую опасность она несет?
Диагноз “генетическая несовместимость” не означает, что пара не может зачать ребенка. Обычно и оплодотворение и имплантация происходит, как при любой естественной беременности. Нарушения обычно проявляются с момента развития эмбриона, а затем в течение беременность прерывается на ранних сроках, обычно в первые 10 недель. Если ситуация повторяется несколько раз может быть поставлен диагноз “привычное невынашивание” и супругов направляют к генетику.
Обычно и оплодотворение и имплантация происходит, как при любой естественной беременности. Нарушения обычно проявляются с момента развития эмбриона, а затем в течение беременность прерывается на ранних сроках, обычно в первые 10 недель. Если ситуация повторяется несколько раз может быть поставлен диагноз “привычное невынашивание” и супругов направляют к генетику.
Почему так происходит?
— Если гены отца и матери отличаются, беременность сохраняется. Организм понимает, что половина генов ребенка от матери, половина от отца. Появляется иммунологическая реакция на сохранение беременности.
— Если гены будущих родителей имеют сходство, то организм включает защитный механизм в ответ на имплантцию эмбриона.
Консультация генетика и генетический анализ
Генетик должен провести тщательный опрос пары, для получения информации об образе жизни, хронических и перенесенных заболеваниях, здоровье родственников и т.
Консультация генетика при планировании беременности обязательно включает в себя расшифровку полученных результатов генетических анализов. Только врач может поставить точный диагноз.
Анализы на генетику при планировании беременности обычно проводятся методом забора крови обоих родителей и занимают до 3 недель.
Как лечится генетическая несовместимость?
При постановке диагноза, врач-генетик вместе с лечащим врачом-репродуктологом или акушер-гинекологом выбирают соответствующую тактику лечения и преодоления трудностей зачатия.
Самый надежный метод достижения успешной беременности и благополучного вынашивания малыша — ЭКО или ИКСИ с поведением дополнительного генетического тестирования эмбриона (ПГТ-А).
В нашем медицинском центре, расположенном в городе Краснодар, гинеколог, генетик и другие врачи проконсультируют будущих родителей по всем интересующим их вопросам. Также, здесь можно будет сдать все необходимые анализы.
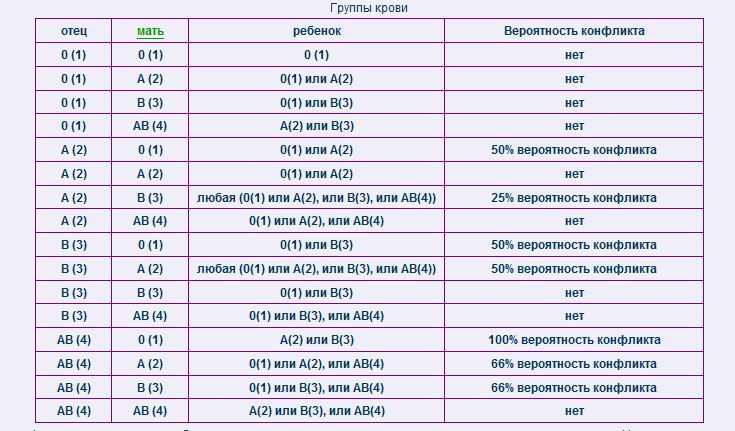
Виды несовместимости партнеров — причины, симптомы, диагностика
Если паре долго не удается зачать ребенка, то проводится ряд исследований, направленных на выяснение причины нарушения фертильности. Но в ряде случаев такие диагностические методы не отличаются информативностью. Напротив, с их помощью удается определить, что ни один из партнеров не имеет существенных проблем. Врачи предполагают, что имеет место неспособность партнеров именно к совместному зачатию.
Здесь требуется сложное прицельное обследование и мужчины, и женщины. После установления причины подобного расстройства подбирается наиболее оптимальная методика решения проблемы. Нередко справиться с ней позволяет проведение консервативного лечения. При его неэффективности прибегают к экстракорпоральному оплодотворению (ЭКО). Раннее обращение за медицинской помощью позволяет своевременно реализовать детородную функцию.
Что такое несовместимость партнеров?
С середины прошлого столетия начали проводиться исследования закономерностей наследственности и генетической изменчивости. Были выдвинуты и первые предположения о причинах неспособности к зачатию некоторых пар или предрасположенности к раннему прерыванию наступившей беременности. Даже при несовершенстве используемых в то время биохимических методик удалось определить часть механизмов, предопределяющих бесплодие пары.
Под несовместимостью понимают патологическое расстройство (конфликт), причем такие нарушения фертильности не единичны. Чаще всего причина кроется в неадекватной реакции иммунной системы на внедрение в женский организм мужского генетического материала. У подобного состояния нет никаких специфических клинических проявлений. В большинстве случаев зачатие и вынашивание беременности бывает успешным с другим партнером.
Чаще всего причина кроется в неадекватной реакции иммунной системы на внедрение в женский организм мужского генетического материала. У подобного состояния нет никаких специфических клинических проявлений. В большинстве случаев зачатие и вынашивание беременности бывает успешным с другим партнером.
Причины несовместимости
Решить проблему с несовместимостью можно только после определения причины патологического состояния. После детального обследования партнеров и проведенного исследования обычно преград для зачатия нет. Чем может быть обусловлена несовместимость женщины и мужчины:
- генетическими аномалиями;
- выработкой антиспермальных антител;
- нарушенным иммунным статусом женского организма;
- близкородственными браками.
Нарушение фертильности может быть спровоцировано и аутоиммунным расстройством. В его основе – повреждение клеток собственного организма иммунной системой в ответ на внедрение чужеродных белков.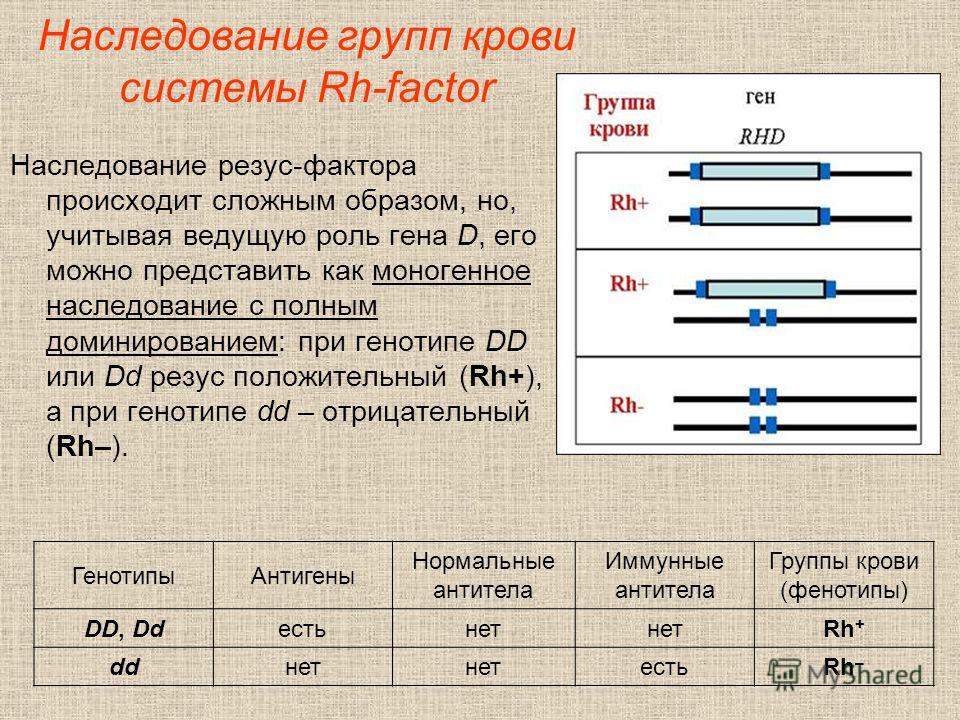
Генетическая несовместимость
Женское и мужское генетическое бесплодие диагностируется приблизительно в 10% клинических случаев расстройств фертильности. Основной причиной становится изменение качественного или количественного состава генов в хромосоме, в которых сосредоточена большая часть наследственной информации. Из-за этого в процессе деления ядра яйцеклетки или сперматозоида образуются дефектные клетки. У них неправильный хромосомный набор, а следовательно, и низкая жизнеспособность. Генетическая несовместимость может быть спровоцирована следующими факторами:
- чрезмерными дозами облучения;
- неблагоприятной экологической обстановкой;
- близкородственными браками;
- генетически обусловленными заболеваниями.
Большое значение имеет и возраст партнеров. По мере естественного старения организма патологически изменяется структура хромосом. У женщин вероятность подобных мутаций особенно высока. Согласно данным клинических исследований, после 35 лет происходит увеличение количества яйцеклеток с генетическими нарушениями. А после 40 лет мутантные ооциты составляют уже 80%. Зачатие и вынашивание беременности в зрелом возрасте бывает вполне успешным, но чаще пара сталкивается с проблемой бесплодия.
А после 40 лет мутантные ооциты составляют уже 80%. Зачатие и вынашивание беременности в зрелом возрасте бывает вполне успешным, но чаще пара сталкивается с проблемой бесплодия.
Тяжелая генетическая поломка становится сигналом для срабатывания механизма естественного отбора. Достаточно быстро или в отсроченном периоде развитие эмбриона, хромосомный набор которого неправилен, перестает развиваться. Беременность замирает или у женщины случается выкидыш. Если это произошло не единожды, то врач может заподозрить закономерность и назначить ряд диагностических исследований.
Наиболее информативны результаты следующих из них:
- кариотипирования;
- экспертной спермограммы;
- FISH-исследования ооцитов и мужских половых клеток.
Установить причину бесплодия конкретной пары позволяет и проведение анализа мутаций в генах, обычно ассоциированных с неспособностью к зачатию и невынашиванием беременности.
Иммунологическая несовместимость
В основе иммунологического (биологического) бесплодия – генетическая тканевая несовместимость. В большинстве случаев привычное прерывание беременности в первом триместре и неудачные имплантации базируются на иммунном отторжении эмбриона или уже начавшего формироваться плода. После того, как произошло зачатие, в норме в женском организме возникает иммуносупрессивное состояние. Иммунная система как бы не обращает внимания на присутствие чужих антигенов в виде примерно половины зародыша. Он благополучно имплантируется к внутренней стенке матки и развивается на протяжении 9 месяцев.
В большинстве случаев привычное прерывание беременности в первом триместре и неудачные имплантации базируются на иммунном отторжении эмбриона или уже начавшего формироваться плода. После того, как произошло зачатие, в норме в женском организме возникает иммуносупрессивное состояние. Иммунная система как бы не обращает внимания на присутствие чужих антигенов в виде примерно половины зародыша. Он благополучно имплантируется к внутренней стенке матки и развивается на протяжении 9 месяцев.
Если же есть несовместимость, то ответ иммунной системы неадекватен. Плодное яйцо считается ею чужеродным материалом, который необходимо отторгнуть. Подобная реакция может быть обусловлена следующими факторами:
- хроническим воспалением слизистой матки;
- эндокринными расстройствами;
- аутоиммунными патологиями.
Оценить местную иммунную защиту эндометрия позволяют данные пайпель-биопсии. В ходе процедуры забирается биологический материал, а затем проводятся иммуногистохимическое и морфологическое исследования.
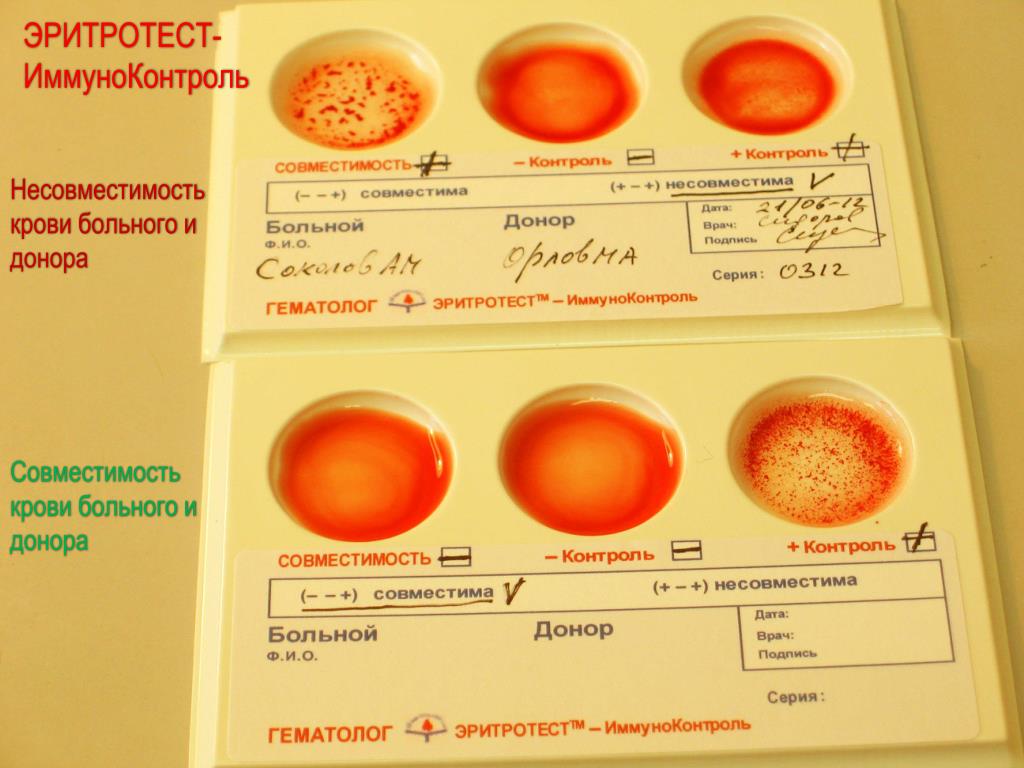
Проверка совместимости пары
Ранее оценить совместимость пары позволяло только выполнение посткоитального теста, направленного на выявление в шеечной (цервикальной) слизи подвижных сперматозоидов. Со временем выяснилось, что его результаты были не столько объективными, сколько субъективными. Сейчас такое исследование почти не проводится из-за низкой диагностической значимости.
В проверке нуждаются пары, ведущие регулярную половую жизнь без использования контрацептивов, если не происходит наступления зачатия и отсутствуют нарушения, которые могли быть выявлены обследованием по стандартному протоколу. Она необходима и при ставших привычными выкидышах, замерших беременностях.
Самые информативные диагностические исследования
Выбор оптимальной для пары методики обследования определяют клиническая ситуация и анамнестические данные. При подозрении на наличие одного из вариантов несовместимости к зачатию выполняются следующие мероприятия:
- определяются антиспермальные антитела у женщины и мужчины исследованием плазмы, спермы или цервикального отделяемого;
- проводятся медико-генетические консультации, цитогенетическое исследование, позволяющее обнаружить отклонения в структуре и числе хромосом.

При низкой информативности данных подобной диагностики, возникших сомнениях о причинах нарушения фертильности детально анализируются ДНК партнеров. Тестирование отличается достаточно высокой стоимостью, но точная причина бесплодия с его помощью будет установлена.
Решение проблемы
Есть несколько методов устранения несовместимости пары к зачатию. Оптимальный вариант терапии будет предложен сразу после выставления диагноза.
- Иммунное бесплодие. Назначается проведение лимфоцитоиммунотерапии для коррекции иммунного ответа женского организма на внедрение чужеродного биологического материала. Повысить степень защиты эмбриона нередко помогает одновременный курсовой прием иммунных препаратов или гормональных средств.
- Генетическое бесплодие. Вспомогательная репродукция выполняется для оценки хромосомного и генного набора эмбриона до его помещения в полость матки. Такой метод терапии позволяет не допустить передачи наследственных патологий.
Если обнаружены антиспермальные антитела у одного из партнеров, то им рекомендована регулярная половая жизнь на протяжении месяца с использованием презервативов. Так удается добиться снижения чрезмерного уровня чувствительности к чужеродным белкам. В комбинации с десенсибилизирующей и иммуносупрессивной терапией подобный метод обычно позволяет устранить проблемы с зачатием.
Так удается добиться снижения чрезмерного уровня чувствительности к чужеродным белкам. В комбинации с десенсибилизирующей и иммуносупрессивной терапией подобный метод обычно позволяет устранить проблемы с зачатием.
Поможет ли ЭКО, если есть несовместимость пары?
Безусловно! Благодаря экстракорпоральному оплодотворению многие столкнувшиеся с несовместимостью пары стали родителями. Нивелирование репродуктологами отрицательного воздействия негативных внутренних факторов приводит к зачатию и благополучному вынашиванию беременности.
В клинике «Чудо Доктор» предоставляются услуги по установлению причины несовместимости пары и терапии разнообразных форм женского и мужского бесплодия. Здесь работают врачи узких специальностей с большим практическим опытом и профессиональным чутьем, подкрепленным огромными теоретическими познаниями. Они ведут пару от первого обращения в клинику до момента успешного родоразрешения.
Автор
Жаранова Елена Васильевна
акушер-гинеколог
Врач высшей категории
Стаж 35 лет
+7 (495) 032-15-21
Генетическая несовместимость широко распространена внутри видов
- Опубликовано:
- Рассел Б. Корбетт-Детиг 1 ,
- Jun Zhou 1 ,
- Эндрю Г. Кларк 2,3 ,
- Дэниел Л. Хартл 1 и
- …
- Julien F. Аэролес 1,2,4
Природа том 504 , страницы 135–137 (2013 г.)Процитировать эту статью
-
20 тыс. обращений
-
129 цитирований
-
66 Альтметрический
-
Сведения о показателях
Субъекты
- Эволюционная генетика
- Генетическое взаимодействие
Abstract
Важность эпистаза — неаддитивных взаимодействий между аллелями — в формировании приспособленности популяции долгое время была спорной темой, чему частично мешало отсутствие эмпирических данных 1,2,3,4 . Традиционно эпистаз выводится на основе независимости генотипических значений между локусами для данного признака. Однако эпистаз для пригодности также должен иметь геномный след 5,6,7 . Чтобы зафиксировать этот сигнал, мы разработали простой подход, основанный на обнаружении искажения соотношения генотипов как признака эпистаза, и мы применяем этот метод к большой панели Drosophila melanogaster рекомбинантных инбредных линий 8,9 . Здесь мы экспериментально подтверждаем, что случаи искажения соотношения генотипов представляют локусы с эпистатическими эффектами приспособленности; по нашим консервативным оценкам, ожидается, что любые два гаплоидных генома в этом исследовании будут содержать 1,15 пары эпистатически взаимодействующих аллелей. Это наблюдение имеет важное значение для генетики видообразования, поскольку оно указывает на то, что исходный материал, обеспечивающий репродуктивную изоляцию, сегрегируется внутри вида одновременно и не обязательно требует, как это предлагается моделью Добжанского-Мюллера, появления несовместимых мутаций, независимо полученных и закрепленных в организме.
Традиционно эпистаз выводится на основе независимости генотипических значений между локусами для данного признака. Однако эпистаз для пригодности также должен иметь геномный след 5,6,7 . Чтобы зафиксировать этот сигнал, мы разработали простой подход, основанный на обнаружении искажения соотношения генотипов как признака эпистаза, и мы применяем этот метод к большой панели Drosophila melanogaster рекомбинантных инбредных линий 8,9 . Здесь мы экспериментально подтверждаем, что случаи искажения соотношения генотипов представляют локусы с эпистатическими эффектами приспособленности; по нашим консервативным оценкам, ожидается, что любые два гаплоидных генома в этом исследовании будут содержать 1,15 пары эпистатически взаимодействующих аллелей. Это наблюдение имеет важное значение для генетики видообразования, поскольку оно указывает на то, что исходный материал, обеспечивающий репродуктивную изоляцию, сегрегируется внутри вида одновременно и не обязательно требует, как это предлагается моделью Добжанского-Мюллера, появления несовместимых мутаций, независимо полученных и закрепленных в организме. аллопатрии. Актуальность нашего результата выходит за рамки видообразования, поскольку он демонстрирует, что эпистаз широко распространен, но часто может оставаться незамеченным из-за отсутствия статистической мощности или отсутствия полногеномного охвата экспериментов.
аллопатрии. Актуальность нашего результата выходит за рамки видообразования, поскольку он демонстрирует, что эпистаз широко распространен, но часто может оставаться незамеченным из-за отсутствия статистической мощности или отсутствия полногеномного охвата экспериментов.
Это предварительный просмотр содержимого подписки, доступ через ваше учреждение
Соответствующие статьи
Статьи открытого доступа со ссылками на эту статью.
-
Протеотоксичность, вызванная нарушенными белковыми комплексами, лежит в основе несовместимости гибридов у дрожжей.
- Кришна Б.С. Свами
- , Синь-И Ли
- … Джун-И Леу
Связь с природой Открытый доступ 29 июля 2022 г.
-
Выявление эпистаза приспособленности в недавно смешанных популяциях с помощью полногеномных данных
- Сюмин Ни
- , Мэнши Чжоу
- … Сяофэн Чжу
Геномика BMC Открытый доступ 11 июля 2020 г.
-
Искажение коэффициента передачи часто встречается в контролируемых скрещиваниях Arabidopsis thaliana.
- Данель К. Сеймур
- , Юнён Че
- … Детлеф Вайгель
Наследственность Открытый доступ 28 июня 2018 г.

Варианты доступа
Подписаться на журнал
Получить полный доступ к журналу на 1 год
199,00 €
всего 3,90 € за выпуск
Подписаться
Расчет налога будет завершен во время оформления заказа.
Купить статью
Получите ограниченный по времени или полный доступ к статье на ReadCube.
32,00 $
Купить
Все цены указаны без учета стоимости.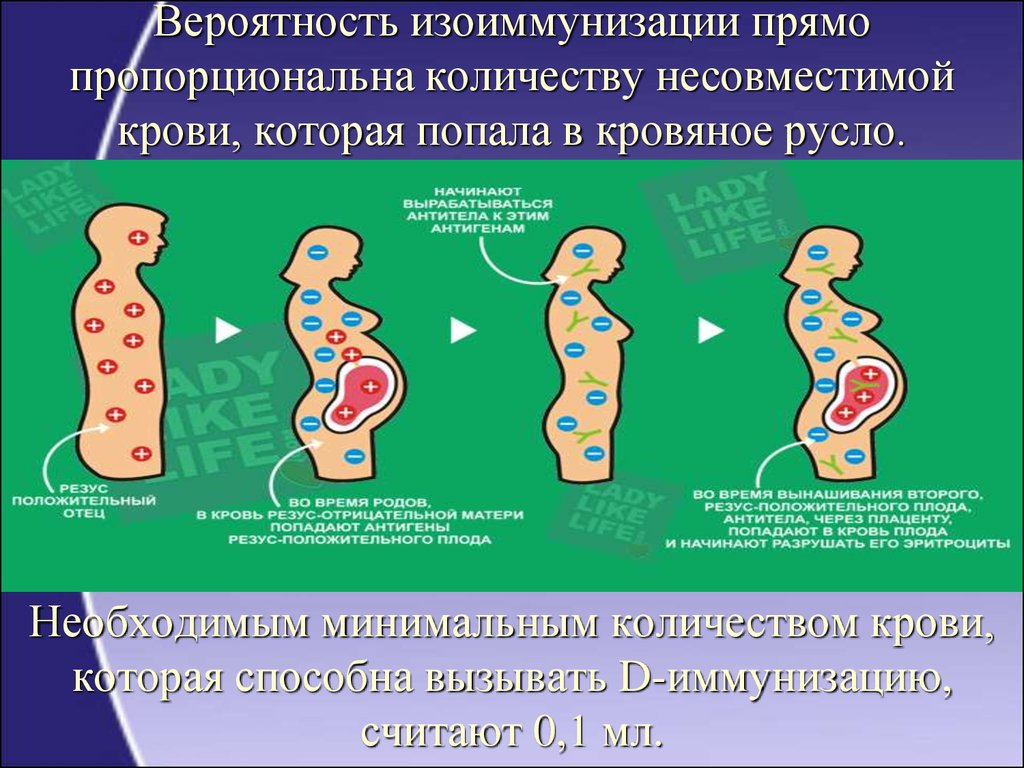
Ссылки
-
Presgraves, D.C. Молекулярно-эволюционная основа формирования видов. Природа Преподобный Жене. 11 , 175–180 (2010)
Статья КАС пабмед Google ученый
-
Койн, Дж. А. и Орр, Х. А. Спецификация (Sinauer Associates, 2004)
Google ученый
-
Каттер А.Д. Полиморфная прелюдия к несовместимости Бейтсона-Добжанского-Мюллера. Тренды Экол. Эвол. 27 , 209–218 (2012)
Статья пабмед Google ученый
-
Carlborg, O.
& Haley, C.S. Эпистаз: слишком часто пренебрегают исследованиями сложных признаков? Природа Преподобный Жене. 5 , 618–625 (2004) Статья КАС пабмед Google ученый
-
Phillips, P.C. Эпистаз — существенная роль взаимодействий генов в структуре и эволюции генетических систем. Природа Преподобный Жене. 9 , 855–867 (2008)
Статья КАС пабмед Google ученый
-
Бомблис, К. и др. Аутоиммунный ответ как механизм синдрома несовместимости типа Добжанского-Мюллера у растений. PLoS Биол. 5 , e236 (2007)
Артикул пабмед ПабМед Центральный КАС Google ученый
-
Payseur, B.A. & Hoekstra, HE. Признаки репродуктивной изоляции в моделях однонуклеотидного разнообразия среди инбредных штаммов домашних мышей.
Генетика 171 , 1905–1916 (2005) Статья КАС пабмед ПабМед Центральный Google ученый
-
Кинг, Э. Г., Макдональд, С. Дж. и Лонг, А. Д. Свойства и возможности Drosophila Synthetic Population Resource для рутинного анализа сложных признаков. Генетика 191 , 935–949 (2012)
Статья пабмед ПабМед Центральный Google ученый
-
Кинг, Э. Г. и др. Генетическое вскрытие модельного сложного признака с использованием ресурса синтетической популяции Drosophila . Рез. генома. 22 , 1558–1566 (2012)
Артикул КАС пабмед ПабМед Центральный Google ученый
-
Zuk, O. et al. Тайна отсутствия наследуемости: генетические взаимодействия создают фантомную наследственность.
Проц. Натл акад. науч. США 109 , 1193–1198 (2012) Статья ОБЪЯВЛЕНИЯ КАС пабмед ПабМед Центральный Google ученый
-
Бикард, Д. и др. Дивергентная эволюция дублированных генов приводит к генетической несовместимости внутри A. thaliana . Наука 323 , 623–626 (2009)
Статья ОБЪЯВЛЕНИЯ КАС пабмед Google ученый
-
Палополи, М. Ф. и Ву, К. И. Генетика гибридного мужского бесплодия между родственными видами дрозофилы : в межвидовых исследованиях выявлена сложная паутина эпистаза. Генетика 138 , 329–341 (1994)
Статья КАС пабмед ПабМед Центральный Google ученый
-
Ву, К.И., Джонсон, Н.А. и Палополи, М.Ф. Правило Холдейна и его наследие: почему так много бесплодных мужчин? Тренды Экол.
Эвол. 11 , 281–284 (1996) Статья КАС пабмед Google ученый
-
Wasbrough, E. R. et al. Протеом-II спермы Drosophila melanogaster (DmSP-II). J. Proteomics 73 , 2171–2185 (2010)
Артикул КАС пабмед Google ученый
-
Докендорф Т.С., Робертсон С.Е., Фолкнер Д.Л. и Йонгенс Т.А. Генетическая характеристика области 44D–45B генома Drosophila melanogaster на основе летального скрининга F2. Мол. Генерал Жене. 263 , 137–143 (2000)
Статья КАС пабмед Google ученый
-
Нетцель-Арнетт, С. и др. Гликозилфосфатидилинозитол-заякоренная сериновая протеаза PRSS21 (тестизин) придает мышиным эпидидимальным сперматозоидам созревание и способность к оплодотворению. Биол.
Воспр. 81 , 921–932 (2009) Статья КАС пабмед ПабМед Центральный Google ученый
-
Касаи, С. и Томита, Т. Специфическая мужская экспрессия цитохрома Р450 (Cyp312a1) в Drosophila melanogaster . Биохим. Биофиз. Рез. коммун. 300 , 894–900 (2003)
Артикул КАС пабмед Google ученый
-
Мейкледжон, К.Д., Монтут, К.Л. и Рэнд, Д.М. Положительный и отрицательный отбор в митохондриальном геноме. Тенденции Жене. 23 , 259–263 (2007)
Статья КАС пабмед Google ученый
-
Ковер, П. X. и др. Многородительское перекрестное скрещивание продвинутого поколения для тонкой карты количественных признаков у Arabidopsis thaliana . Генетика PLoS. 5 , e1000551 (2009)
Артикул пабмед ПабМед Центральный КАС Google ученый
-
McMullen, M.
D. et al. Генетические свойства картографической популяции вложенных ассоциаций кукурузы. Наука 325 , 737–740 (2009) Артикул ОБЪЯВЛЕНИЯ КАС пабмед Google ученый
-
Hill, W.G., Goddard, M.E. & Visscher, P.M. Данные и теория указывают в основном на аддитивную генетическую изменчивость сложных признаков. Генетика PLoS. 4 , e1000008 (2008)
Артикул пабмед ПабМед Центральный КАС Google ученый
-
Добжанский Т. Генетика и происхождение видов (Columbia Univ. Press, 1937)
Google ученый
-
Орр, Х. А. и Турелли, М. Эволюция постзиготической изоляции: накопление несовместимости Добжанского-Мюллера. Evolution 55 , 1085–1094 (2001)
Артикул КАС пабмед Google ученый
-
Пресгрейвс, Д.
К. и Стефан, В. Всепроникающая адаптивная эволюция среди взаимодействующих Ген гибридной нежизнеспособности дрозофилы , Nup96 . Мол. биол. Эвол. 24 , 306–314 (2007) Статья КАС пабмед Google ученый
-
Тао Ю. и др. Генетическое вскрытие гибридной несовместимости между Drosophila simulans и D. mauritiana . I. Дифференциальное накопление влияния гибридной мужской стерильности на X и аутосомы. Генетика 164 , 1383–1397 (2003)
Статья КАС пабмед ПабМед Центральный Google ученый
-
Фитцпатрик Б.М. Гибридная дисфункция: популяционно-генетические и количественные генетические перспективы. утра. Нац. 171 , 491–498 (2008)
Статья пабмед Google ученый
-
Демут, Дж.
П. и Уэйд, М. Дж. О теоретической и эмпирической основе для изучения генетических взаимодействий внутри и между видами. утра. Нац. 165 , 524–536 (2005) Статья пабмед Google ученый
-
Рид Л.К. и Маркоу Т.А. Ранние события видообразования: полиморфизм гибридной мужской стерильности у Drosophila . Проц. Натл акад. науч. США 101 , 9009–9012 (2004)
Статья ОБЪЯВЛЕНИЯ КАС пабмед ПабМед Центральный Google ученый
-
Чеверуд, Дж. М. и Рутман, Э. Дж. Эпистаз и его вклад в компоненты генетической изменчивости. Генетика 139 , 1455–1461 (1995)
Статья КАС пабмед ПабМед Центральный Google ученый
-
King, E.G. et al. Генетическое вскрытие модельного сложного признака с использованием ресурса синтетической популяции дрозофилы.
Рез. генома. 22 , 1558–1566 (2012) Артикул КАС пабмед ПабМед Центральный Google ученый
-
Кинг, Э. Г., Макдональд, С. Дж. и Лонг, А. Д. Свойства и возможности ресурса синтетической популяции дрозофилы для рутинного анализа сложных признаков. Генетика 191 , 935–949 (2012)
Статья пабмед ПабМед Центральный Google ученый
-
Baird, N. A. et al. Быстрое обнаружение SNP и генетическое картирование с использованием секвенированных маркеров RAD. PLoS ONE 3 , e3376 (2008)
Артикул ОБЪЯВЛЕНИЯ пабмед ПабМед Центральный КАС Google ученый
-
Акерманн, М. и Бейер, А. Систематическое обнаружение эпистатических взаимодействий на основе частот пар аллелей. Генетика PLoS.
8 , e1002463 (2012) Артикул КАС пабмед ПабМед Центральный Google ученый
-
Бенджамини Ю. и Хохберг Ю. Контроль уровня ложных открытий: практичный и мощный подход к множественному тестированию. JR Stat. соц. B 57 , 289–300 (1995)
MathSciNet МАТЕМАТИКА Google ученый
-
Phillips, P.C. Эпистаз — существенная роль взаимодействий генов в структуре и эволюции генетических систем. Природа Преподобный Жене. 9 , 855–867 (2008)
Артикул КАС пабмед Google ученый
-
Чеверуд, Дж. М. и Рутман, Э. Дж. Эпистаз и его вклад в компоненты генетической изменчивости. Генетика 139 , 1455–1461 (1995)
Статья КАС пабмед ПабМед Центральный Google ученый
-
Adams, M.
D. et al. Последовательность генома Drosophila melanogaster . Наука 287 , 2185–2195 (2000) Статья пабмед Google ученый
-
Лунтер, Г. и Гудсон, М. Стэмпи: Статистический алгоритм для чувствительного и быстрого картирования прочтений последовательности Illumina. Рез. генома. 21 , 936–939 (2011)
Статья КАС пабмед ПабМед Центральный Google ученый
-
Ли, Х. и Дурбин, Р. Быстрое и точное выравнивание коротких считываний с преобразованием Берроуза-Уилера. Биоинформатика 25 , 1754–1760 (2009)
Статья КАС пабмед ПабМед Центральный Google ученый
-
Де Присто, М. А. и др. Основа для обнаружения вариаций и генотипирования с использованием данных секвенирования ДНК следующего поколения.
Природа Жене. 43 , 491–498 (2011) Статья КАС пабмед Google ученый
-
Cingolani, P. et al. Программа для аннотирования и прогнозирования эффектов однонуклеотидных полиморфизмов, SnpEff: SNPs в геноме Drosophila melanogaster штамм w 1118 ; изо-2; изо-3. Fly (Остин) 6 , 80–82 (2012)
Статья КАС Google ученый
-
McMullen, M.D. et al. Генетические свойства картографической популяции вложенных ассоциаций кукурузы. Наука 325 , 737–740 (2009)
Статья ОБЪЯВЛЕНИЯ КАС пабмед Google ученый
-
Kover, P. X. et al. Многородительское перекрестное скрещивание в старшем поколении для тонкой карты количественных признаков у Arabidopsis thaliana .
Генетика PLoS. 5 , e1000551 (2009) Артикул пабмед ПабМед Центральный КАС Google ученый
 & Haley, C.S. Эпистаз: слишком часто пренебрегают исследованиями сложных признаков? Природа Преподобный Жене. 5 , 618–625 (2004)
& Haley, C.S. Эпистаз: слишком часто пренебрегают исследованиями сложных признаков? Природа Преподобный Жене. 5 , 618–625 (2004) 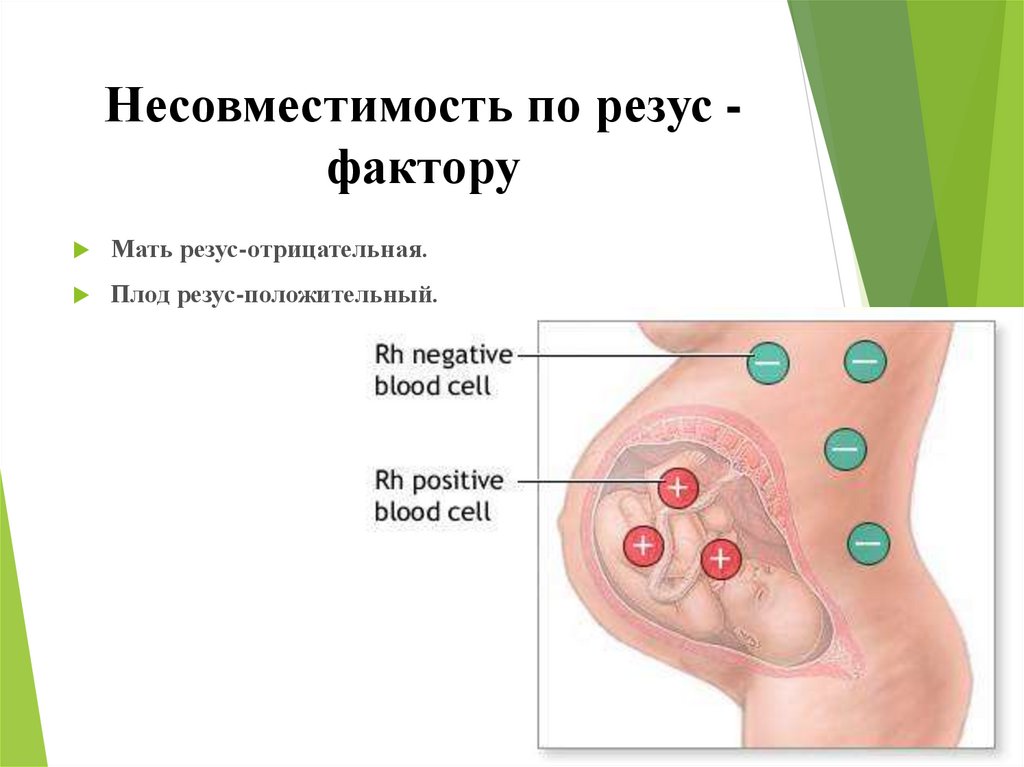 Генетика 171 , 1905–1916 (2005)
Генетика 171 , 1905–1916 (2005)  Проц. Натл акад. науч. США 109 , 1193–1198 (2012)
Проц. Натл акад. науч. США 109 , 1193–1198 (2012)  Эвол. 11 , 281–284 (1996)
Эвол. 11 , 281–284 (1996) 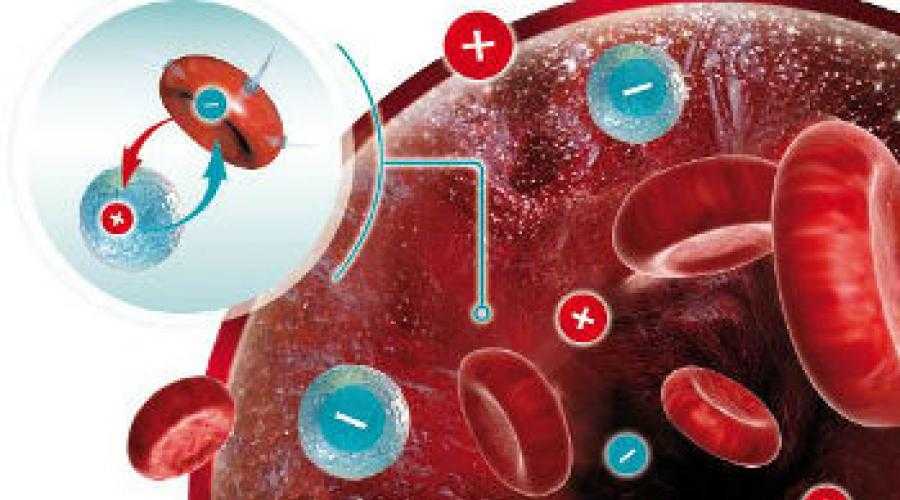 Воспр. 81 , 921–932 (2009)
Воспр. 81 , 921–932 (2009) 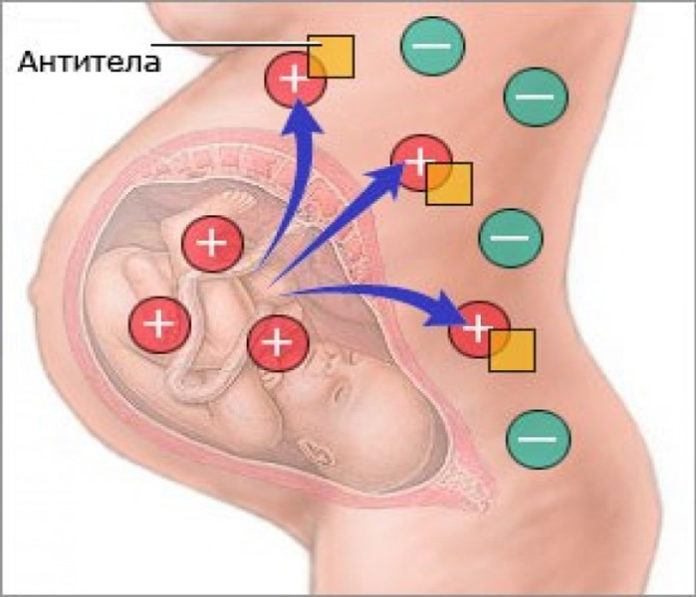 D. et al. Генетические свойства картографической популяции вложенных ассоциаций кукурузы. Наука 325 , 737–740 (2009)
D. et al. Генетические свойства картографической популяции вложенных ассоциаций кукурузы. Наука 325 , 737–740 (2009)  П. и Уэйд, М. Дж. О теоретической и эмпирической основе для изучения генетических взаимодействий внутри и между видами. утра. Нац. 165 , 524–536 (2005)
П. и Уэйд, М. Дж. О теоретической и эмпирической основе для изучения генетических взаимодействий внутри и между видами. утра. Нац. 165 , 524–536 (2005)  Рез. генома. 22 , 1558–1566 (2012)
Рез. генома. 22 , 1558–1566 (2012) 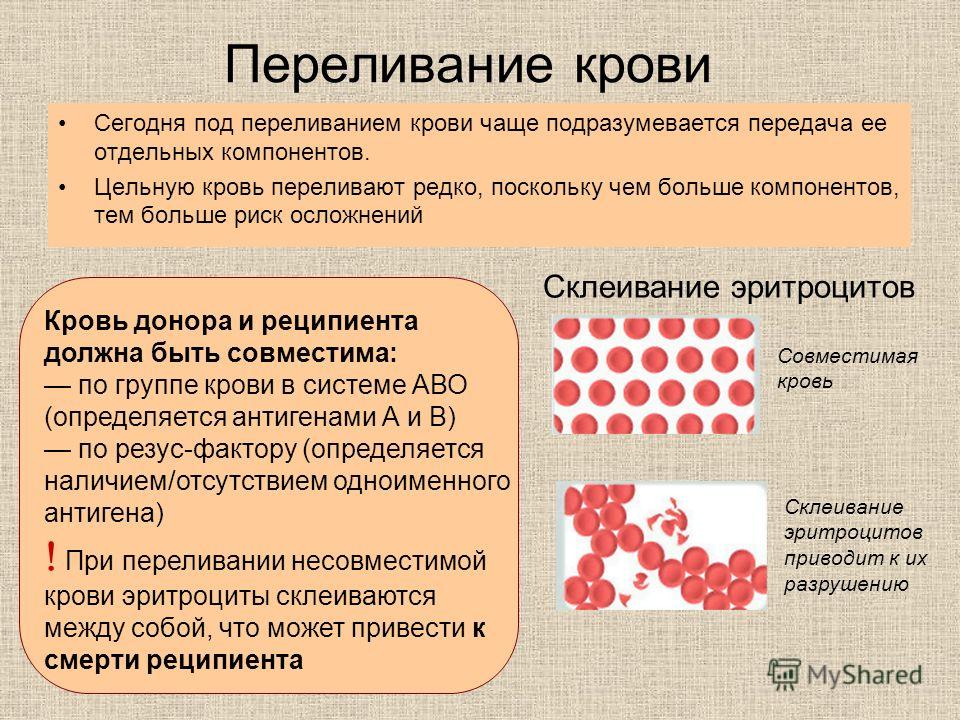 8 , e1002463 (2012)
8 , e1002463 (2012)  Природа Жене. 43 , 491–498 (2011)
Природа Жене. 43 , 491–498 (2011)  Генетика PLoS. 5 , e1000551 (2009)
Генетика PLoS. 5 , e1000551 (2009) Ссылки на скачивание
Благодарности
Мы благодарны Т. Лонгу, С. Макдональду и Э. Кингу за создание DSPR и предоставление нам RIL и штаммов-основателей. Мы благодарим К. Джонса, Б. де Биворта, Т. Сактона, С. Д. Кохера, Дж. Гренье и Н.Е. Солтису за комментарии и обсуждения. Мы благодарим X. Ши за техническую помощь. Эта работа была поддержана грантами: NIH GM065169 и GM084236 для DLH, HD059060 для AGC и Гарвардского общества стипендиатов и фондов Гарварда Милтона для JFA. РБК-Д был поддержан стипендией Гарвардской премии.
Информация об авторе
Авторы и организации
-
Факультет органической и эволюционной биологии, Гарвардский университет, Кембридж, 02138, Массачусетс, США
Рассел Б. Корбетт-Детиг, Джулиан Ф. Эйролес
-
Кафедра биологической статистики и вычислительной биологии, Корнельский университет, Итака, 14853, Нью-Йорк, США
Эндрю Г.
Кларк и Жюльен Ф. Эйролес -
Кафедра молекулярной биологии и генетики, Корнельский университет, Итака, 14853, Нью-Йорк, США
Эндрю Г. Кларк
-
Гарвардское общество стипендиатов, Гарвардский университет, Кембридж, 02138, Массачусетс, США
Жюльен Ф. Эйролес
 Кларк и Жюльен Ф. Эйролес
Кларк и Жюльен Ф. Эйролес Авторы
- Russell B. Corbett-Detig
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Scholar
- Jun Zhou
Посмотреть публикации автора
Вы также можете искать этого автора в PubMed Google Scholar
- Andrew G. Clark
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Scholar
- Daniel L. Hartl
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Scholar
- Julien F. Ayroles
Посмотреть публикации автора
Вы также можете искать этого автора в PubMed Google Scholar
 Ayroles
Ayroles Contributions
J.F.A. задумал идею проекта, R.B.C.-D. и Дж.Ф.А. задумал и разработал эксперименты и анализы. РБК-Д и Дж.Ф.А. проводил биоинформатический и статистический анализы; РБК-Д., J.F.A. и Дж.З. проведенные эксперименты; Дж.З. выполнял молекулярную работу; А.Г.К. и Д.Л.Х. давал аналитические и концептуальные советы на протяжении всего проекта.
Автор, ответственный за переписку
Переписка с Жюльен Ф. Эйролес.
Заявление об этике
Конкурирующие интересы
Авторы не заявляют об отсутствии конкурирующих финансовых интересов.
Дополнительная информация
Весь код, использованный и сгенерированный для этого исследования, предоставляется по запросу.
Рисунки и таблицы с расширенными данными
Расширенные данные Рис. 1 Описание DSPR и схемы проверки.

a , Географическое распространение штаммов-основателей DSPR (оранжевый, панель A; красный, панель B). b , Конструирование рекомбинантных инбредных линий. Для каждой панели все штаммы-основатели были скрещены по круговой схеме (линия 1 ♀ × линия 2 ♂, линия 2 ♀ × линия 3 ♂,…, линия 8 ♀ × линия 1 ♂) для получения F 1 s, и Затем особям F 1 было позволено свободно спариваться, чтобы произвести популяцию F 2 . В каждой панели A и B эти популяции F 2 были разделены на две независимые популяции для создания панелей A1, A2 и B1, B2. Каждому было позволено свободно рекомбинировать в течение 50 поколений в очень большой популяции. После 50 поколений для каждой повторной панели около 400 изоженских линий были инбредированы в течение 25 поколений для создания 4 панелей RIL, используемых в этом исследовании. c , Схема скрещивания, используемая для проверки эпистатических эффектов. Пара основателей, разделяющих несовместимые аллели, была выбрана и скрещена для получения F 1 s; затем мы скрестили потомство F 1 , чтобы получить большую популяцию F 2 , разделив все возможные комбинации аллелей между аллелями в локусах 1 и 2. Затем мы подсчитали потомство, полученное от каждой пары путем скрещивания большого количества F 2 . s, которые позже были генотипированы в сайтах, близких к предсказанным взаимодействующим локусам.
Затем мы подсчитали потомство, полученное от каждой пары путем скрещивания большого количества F 2 . s, которые позже были генотипированы в сайтах, близких к предсказанным взаимодействующим локусам.
Расширенные данные Рис. 2. Анализ основных компонентов всех трех панелей DSPR RIL.
Зеленый, панель A-2; синий, панель Б-1; и красный, панель B-2. Никаких доказательств структуры населения не показано.
Расширенные данные Рисунок 3 Распределение D’ для значительного GRD.
Данные отображаются на панелях DSPR. На оси x D’ является мерой неравновесия между взаимодействующими аллелями. Красная кривая соответствует плавной подгонке кривой с использованием непараметрической оценки плотности. Коробчатая диаграмма с выбросами представлена над гистограммой (ромбы представляют собой среднее и 95% ДИ, край прямоугольника представляет процентиль 25% и 75%, вертикальная черта внутри медианы, точки возможны выбросы, а красная скобка представляет собой наименьшую длину, содержащую 50% данных).
Расширенные данные Рис. 4 График эпистаза для каждого подтвержденного случая GRD.
На осях и расположены измерения продуктивности, соответствующие каждому генотипическому классу по обеим хромосомам. Оси x соответствуют генотипам на одной из хромосом, другой генотип представлен цветом, указанным внутри графика (например, генотип AA , bb на панели a находится в левом нижнем углу, где AA читается по оси x и bb по синему цвету). a , GRD между хромосомами 2R и 3R (помечены SNP 2R:4806926, по оси X и 3R:5870973, цветные линии) показывает сильный отрицательный эпистаз из-за низкой приспособленности генотипа aa;bb . Генетический эффект от добавки к добавке равен -13,75 (в смысле ссылок 5 и 29).). b , GRD между хромосомами 3L и X (помечены SNP 3L: 11510853, по оси X и X: 16483812, цветные линии) также показывает отрицательный эпистаз. Здесь пошаговый генетический эффект равен -5,94.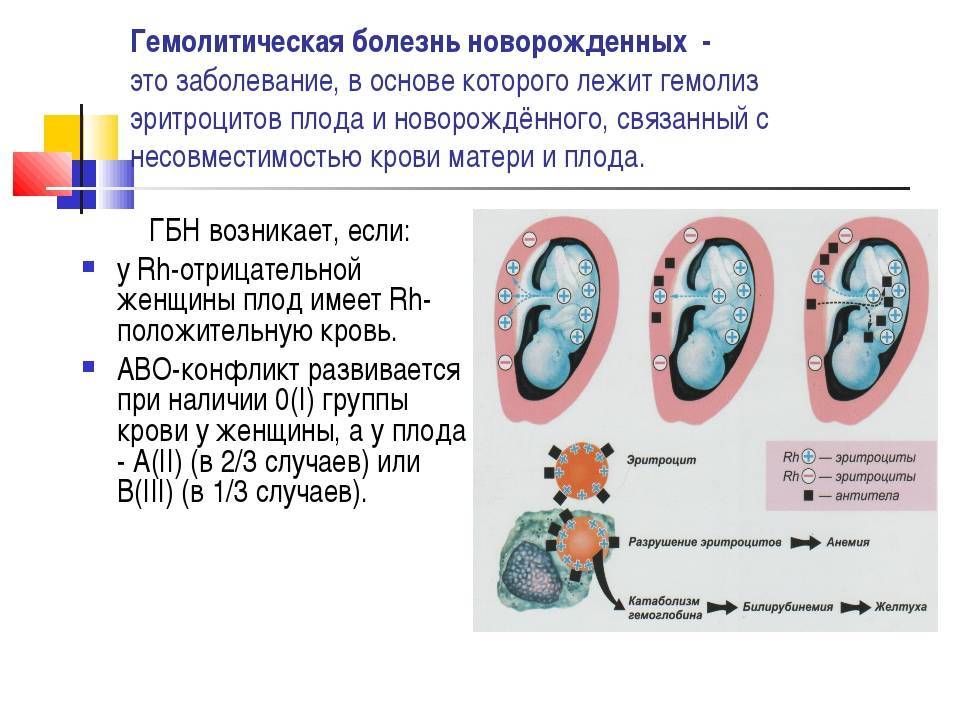
Расширенные данные Рисунок 5. Накопление постзиготической репродуктивной изоляции во времени (обратите внимание на логарифмическую шкалу на осях).
Приблизительное время расхождения обычно изучаемых видов Drosophila обозначено зелеными кружками, а красный кружок указывает разумное ожидание времени расхождения запасов, использованных для основания DSPR (~10 000 лет). Горизонтальная красная область указывает на очень приблизительный «порог видообразования» и указывает на то, что многие обычно изучаемые пары видов значительно превышают этот порог.
Расширенная таблица данных 1 Список всех значимых межхромосомных GRD, идентифицированных в DSPRПолная таблица
Расширенная таблица данных 2 Список значимых межхромосомных GRD в панели Arabidopsis MAGIC и панели NAM кукурузыПолная Таблица размеров
Слайды PowerPoint
Слайд PowerPoint для рисунка 1
Слайд PowerPoint для рисунка 2
Слайд PowerPoint для рисунка 3
Права и разрешения
Перепечатка и разрешения
Об этой статье
Эта статья цитируется
-
Протеотоксичность, вызванная нарушенными белковыми комплексами, лежит в основе несовместимости гибридов у дрожжей.
- Кришна Б.С. Свами
- Синь-И Ли
- Джун-И Леу
Nature Communications (2022)
-
Пространственно-временная динамика взаимодействующих генетических несовместимостей. Часть I: случай сложенных недоминантных клинов
- Матье Альфаро
- Квентин Гриетт
- Бенуа Сарельс
Журнал математической биологии (2022)
-
Выявление эпистаза приспособленности в недавно смешанных популяциях с помощью полногеномных данных
- Сюмин Ни
- Мэнши Чжоу
- Сяофэн Чжу
BMC Genomics (2020)
-
Периодическое нарушение и восстановление баланса сегрегационных искажений в геномах: битва за преимущество передачи
- Фан Ся
- Идан Оуян
АБИОТЕК (2020)
-
Мейотический драйв X-хромосомы у Drosophila simulans: подход с использованием QTL раскрывает сложный полигенный детерминизм подавления парижского драйва
- Сесиль Курре
- Пьер Р. Жерар
- Катрин Моншан-Моро
Наследственность (2019)

 Жерар
Жерар Комментарии
Отправляя комментарий, вы соглашаетесь соблюдать наши Условия и Правила сообщества. Если вы обнаружите что-то оскорбительное или не соответствующее нашим условиям или правилам, отметьте это как неприемлемое.
Генетическая несовместимость приводит к выбору партнера у осы-паразита | Границы зоологии
- Краткий отчет
- Открытый доступ
- Опубликовано:
- Andra Thiel 1 ,
- Anne C Weeda 1 ,
- Jetske G de Boer 2,3 &
Границы зоологии том 10 , Номер статьи: 43 (2013) Процитировать эту статью
-
5196 Доступ
-
12 цитирований
-
Сведения о показателях
Abstract
Introduction
Аллельная несовместимость между особями одного и того же вида должна определять выбор партнера на основе генетического состава обоих партнеров в локусах, влияющих на приспособленность потомства. Как следствие, выбор партнера может быть важной движущей силой аллельного разнообразия. Система комплементарного определения пола (CSD) отвечает за внутривидовую аллельную несовместимость у многих видов муравьев, пчел и ос. Таким образом, CSD может способствовать диссортативному спариванию и в этом напоминает MHC иммунной системы позвоночных или систему самонесовместимости (SI) высших растений.
Как следствие, выбор партнера может быть важной движущей силой аллельного разнообразия. Система комплементарного определения пола (CSD) отвечает за внутривидовую аллельную несовместимость у многих видов муравьев, пчел и ос. Таким образом, CSD может способствовать диссортативному спариванию и в этом напоминает MHC иммунной системы позвоночных или систему самонесовместимости (SI) высших растений.
Результаты
Здесь мы показываем, что у моногамной паразитической осы Bracon brevicornis (Wesmael) самки способны отвергать партнеров с несовместимыми аллелями. Принуждение самок к принятию изначально отвергнутых партнеров приводило к искажению соотношения полов и частичному бесплодию потомства.
Выводы
CSD-дезассортативное спаривание в нашем эксперименте происходило независимо от распознавания родственников и предотвращения инбридинга. Последствия выбора партнера для пригодности можно наблюдать непосредственно, на них не влияют воздействия окружающей среды, и они более серьезны, чем в сопоставимых системах (SI или MHC), как для отдельных людей, так и для популяции. Таким образом, наши результаты демонстрируют большой потенциал выбора самками для поддержания высокой приспособленности потомства у этого вида.
Таким образом, наши результаты демонстрируют большой потенциал выбора самками для поддержания высокой приспособленности потомства у этого вида.
Введение
Выбор партнера, неслучайный выбор партнера, чрезвычайно распространен среди животных, размножающихся половым путем. Иногда накапливаются прямые выгоды, такие как ресурсы, но часто самки «покупают» генетические выгоды, чтобы повысить приспособленность своего потомства [1]. Выбор партнера может быть основан на «хороших генах», указывающих на наследуемое качество партнеров, или на «генетической совместимости» [2, 3]. Последняя концепция подразумевает, что выгоды, получаемые от оптимального выбора партнера, зависят от генетического состава обоих партнеров в локусах, влияющих на приспособленность потомства. Некоторые из наиболее изученных примеров выбора партнера на основе генетической совместимости касаются главного комплекса гистосовместимости (MHC) у позвоночных. Разнообразие локусов MHC может усиливать иммунитет против паразитов и патогенов (например, [4]), а предпочтение MHC-несходных партнеров существует у различных животных, таких как рыбы, птицы и млекопитающие, включая людей (например, [5–7]). Выбор партнера обсуждается как важная движущая сила аллельного разнообразия внутри вида [2, 7], даже несмотря на то, что доказательства часто противоречивы в разных исследованиях [6-10]. Это может быть связано с тем, что преимущества пригодности MHC-дезассортативного спаривания зависят от факторов окружающей среды, например. воздействие паразитов и, таким образом, может различаться у разных видов и в зависимости от экспериментальных условий [11, 12].
Выбор партнера обсуждается как важная движущая сила аллельного разнообразия внутри вида [2, 7], даже несмотря на то, что доказательства часто противоречивы в разных исследованиях [6-10]. Это может быть связано с тем, что преимущества пригодности MHC-дезассортативного спаривания зависят от факторов окружающей среды, например. воздействие паразитов и, таким образом, может различаться у разных видов и в зависимости от экспериментальных условий [11, 12].
Хотя генетическая совместимость может улучшать приспособленность позвоночных, она необходима для развития потомства у растений с системой самонесовместимости (SI) [13]. Генетическая совместимость также имеет решающее значение для перепончатокрылых насекомых, демонстрирующих либо однолокусную, либо множественную комплементарную детерминацию пола (sl-CSD или ml-CSD соответственно). У этих видов гетерозиготность по половому локусу (или локусам) напрямую связана с приспособленностью потомства, поскольку она инициирует путь развития самок в оплодотворенных диплоидных яйцах [14]: сыновья обычно развиваются из неоплодотворенных яиц и гаплоидны.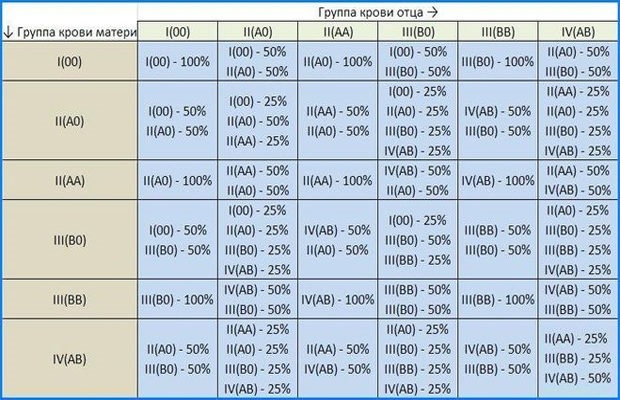 Однако диплоидные самцы образуются, когда яйцеклетки оплодотворяются сперматозоидами с соответствующими половыми аллелями, так что они становятся гомозиготными по половому локусу (sl-CSD, рисунок 1) или половым локусам (ml-CSD). У большинства видов диплоидные самцы нежизнеспособны в развитии или практически бесплодны [15, 16]. Поскольку стерильные диплоидные самцы производятся за счет фертильных самок, последствия спаривания партнера с совпадающим половым аллелем могут быть серьезными.
Однако диплоидные самцы образуются, когда яйцеклетки оплодотворяются сперматозоидами с соответствующими половыми аллелями, так что они становятся гомозиготными по половому локусу (sl-CSD, рисунок 1) или половым локусам (ml-CSD). У большинства видов диплоидные самцы нежизнеспособны в развитии или практически бесплодны [15, 16]. Поскольку стерильные диплоидные самцы производятся за счет фертильных самок, последствия спаривания партнера с совпадающим половым аллелем могут быть серьезными.
Bracon brevicornis репродуктивная биология. Соответствующие спаривания в родительском поколении P приводят к появлению трех видов потомства: диплоидные гомозиготные самцы (F1 слева; одинаковые цветовые полосы на хромосомах), диплоидные гетерозиготные самки (F1 в центре; разные цветные полосы) или гаплоидные гемизиготные самцы из неоплодотворенных яиц (F1 справа).
Изображение в полный размер
Результаты и обсуждение
Оса-паразит Bracon ( Habrobracon ) brevicornis Wesmael (Hymenoptera: Braconidae) (рис. 2) представляет собой превосходную систему для оценки гипотезы о том, что генетическая несовместимость определяет выбор партнера, поскольку: выбор партнера является чисто генетическим, б) самки обычно спариваются только один раз [Weeda & Thiel, неопубликованное наблюдение] и в) генетическая несовместимость приводит к непосредственно измеримым последствиям приспособленности через производство диплоидных самцов [17, 18].
2) представляет собой превосходную систему для оценки гипотезы о том, что генетическая несовместимость определяет выбор партнера, поскольку: выбор партнера является чисто генетическим, б) самки обычно спариваются только один раз [Weeda & Thiel, неопубликованное наблюдение] и в) генетическая несовместимость приводит к непосредственно измеримым последствиям приспособленности через производство диплоидных самцов [17, 18].
Бракон короткорогий габитус. Самка с вставленным яйцекладом сидит на парализованном хозяине (фото: Нильс Линек).
Изображение в полный размер
Мы использовали две популяции, которые, как мы предполагаем, частично перекрываются по присутствующим половым аллелям. Отдельным самкам одной популяции предлагался самец из другой популяции и наоборот, и самка могла либо принять этого самца («принятое» спаривание), либо отказаться от него. Если происходило отторжение, мы заставляли самку принять вторую попытку спаривания того же самца, охлаждая ее на льду («отвергнутое» спаривание). От «отвергнутых» спариваний значительно более высокая доля оплодотворенных яиц развивалась как диплоидные сыновья за счет дочерей (Χ 2 df=1, n=20 = 13,1, P < 0,001; распределение ошибок (ED) = биномиальное, рис. 3). Генетическое совпадение (таблица 1) происходило значительно чаще при «отказных», чем при «принятых» вязках (точный критерий Фишера, односторонний P = 0,009). Однако, поскольку не все «отклоненные» спаривания были совмещены, у самок могли быть дополнительные критерии выбора партнера. При поиске альтернативных объяснений повышенной продукции диплоидных самцов в «отверженных» спаривании мы обнаружили, что ни плодовитость самок (Χ 2 1,20 = 0,007, P = 0,93, табл. 1) ни смертность потомства (Χ 2 1,20 = 1,02, P = 0,31) ос-со- режимы.
От «отвергнутых» спариваний значительно более высокая доля оплодотворенных яиц развивалась как диплоидные сыновья за счет дочерей (Χ 2 df=1, n=20 = 13,1, P < 0,001; распределение ошибок (ED) = биномиальное, рис. 3). Генетическое совпадение (таблица 1) происходило значительно чаще при «отказных», чем при «принятых» вязках (точный критерий Фишера, односторонний P = 0,009). Однако, поскольку не все «отклоненные» спаривания были совмещены, у самок могли быть дополнительные критерии выбора партнера. При поиске альтернативных объяснений повышенной продукции диплоидных самцов в «отверженных» спаривании мы обнаружили, что ни плодовитость самок (Χ 2 1,20 = 0,007, P = 0,93, табл. 1) ни смертность потомства (Χ 2 1,20 = 1,02, P = 0,31) ос-со- режимы.
Последствия выбора партнера. Доля (± SE) всего полученного диплоидного потомства мужского пола и, следовательно, дорогостоящего (*** P < 0,001).
Полноразмерное изображение
Таблица 1 Количество произведенных яиц, гаплоидных самцов (HM), диплоидных самцов (DM), самок (F), неопределенных потомков мужского пола (UM) и вероятность того, что случайное спаривание не привело к образованию диплоидных самцов (вероятность совпадения, рассчитывается как 0,5 (n диплоидное потомство) )Полноразмерная таблица
Несмотря на то, что в течение короткого времени охлаждались только самки от «отвергнутого» спаривания, влияние охлаждения на долю диплоидных самцов, произведенных самками с «отвергнутым» спариванием, маловероятно. 1) Охлаждение произошло в момент времени, который не коррелировал ни с производством яйцеклеток, ни с хранением спермы, ни с оплодотворением. 2) Контрольные самки, которые были спарены без охлаждения с одним из своих сыновей и, таким образом, имели гарантированное спаривание, произвели равное количество диплоидных самцов (X 2 1,15 = 0,14, P = 0,71, таблица 1).
Таким образом, самки продемонстрировали способность снижать производство диплоидных самцов примерно до 1/4 того, что можно было ожидать при случайном спаривании в нашем эксперименте. Так как диплоидные сыновья производятся за счет фертильных дочерей и фактически бесплодны у этого вида [Thiel & Weeda, неопубликованное наблюдение], это ясно демонстрирует избирательное преимущество выбора партнера, обусловленное генетической несовместимостью.
Выводы
Наши результаты показывают, что у насекомого с sl-CSD распознавание специфических аллелей может происходить даже среди неродственных партнеров по спариванию. Таким образом, CSD способствует ауткроссингу, аналогичному системе self-incompatibility (SI), обнаруженной у растений [12], и системе MHC у позвоночных [18]. Тем не менее последствия приспособленности аллельного соответствия более серьезны при CSD, чем при MHC или SI, потому что гомозиготность по локусу CSD неизменно увеличивает постзиготную смертность и производство бесплодного потомства [19].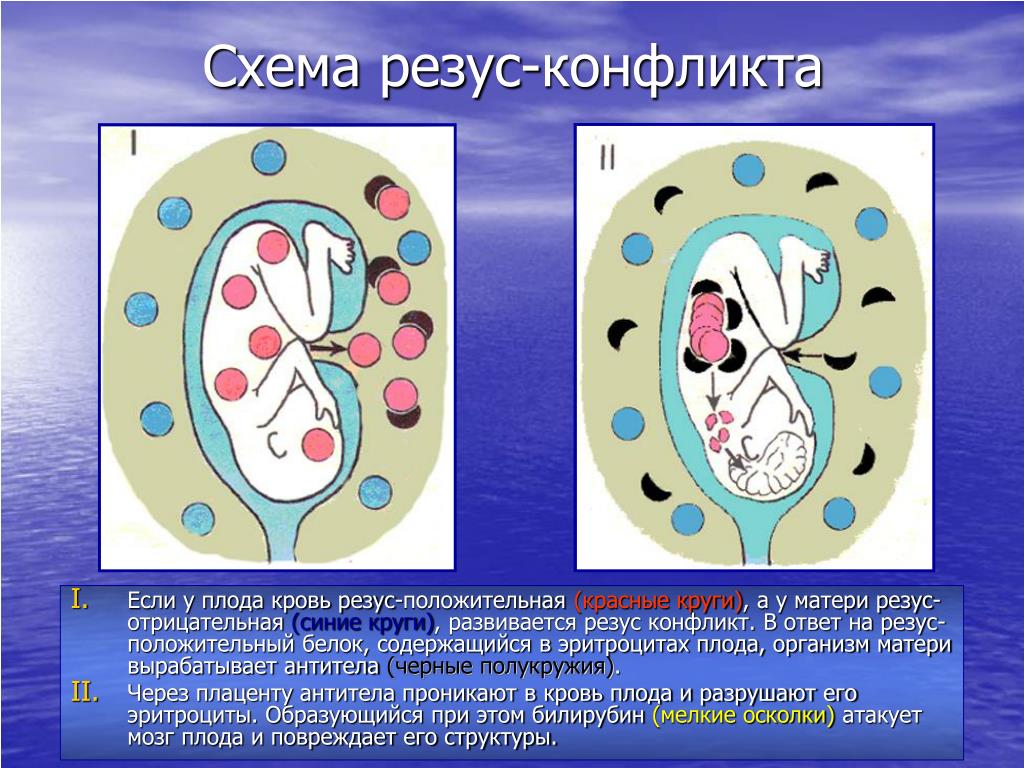 ]. Распознавание родственников по обонятельным сигналам было описано у нескольких видов насекомых (например, [20–22]), и предотвращение инбридинга, безусловно, является важной мерой в снижении вероятности, с которой может произойти согласованное спаривание [16]. Однако в популяциях с узкими местами даже неродственные особи могут иметь общий аллель, определяющий пол. Как следствие, теоретические модели предсказывают быстрое сокращение эффективного размера популяции и высокую вероятность вымирания [23, 24]. Таким образом, выбор самок на основе «генетической совместимости», как описано в нашем исследовании, можно рассматривать как важный механизм повышения выживаемости популяции, а также индивидуальной приспособленности ос.
]. Распознавание родственников по обонятельным сигналам было описано у нескольких видов насекомых (например, [20–22]), и предотвращение инбридинга, безусловно, является важной мерой в снижении вероятности, с которой может произойти согласованное спаривание [16]. Однако в популяциях с узкими местами даже неродственные особи могут иметь общий аллель, определяющий пол. Как следствие, теоретические модели предсказывают быстрое сокращение эффективного размера популяции и высокую вероятность вымирания [23, 24]. Таким образом, выбор самок на основе «генетической совместимости», как описано в нашем исследовании, можно рассматривать как важный механизм повышения выживаемости популяции, а также индивидуальной приспособленности ос.
Материалы и методы
Использованы образцы из двух лабораторных популяций стадных личиночных эктопаразитоидов Bracon ( Habrobracon ) brevicornis Wesmael (Hymenoptera: Braconidae) из сборов 2006 и (2006) двух разных лет тот же полевой участок недалеко от Лейпцига, Германия. Во время эксперимента две популяции были разделены примерно на 60 поколений, и, таким образом, их соответствующие представители, вероятно, перекрывались по присутствующим аллелям, определяющим пол, но не представляли собой близкого родства. Личинки Ephestia kuehniella Zeller (Lepidoptera: Pyralidae) служили хозяевами. Эксперименты и выращивание проводили при 25°C, 55% отн. вл. и 16:8 ч свет:темно.
Во время эксперимента две популяции были разделены примерно на 60 поколений, и, таким образом, их соответствующие представители, вероятно, перекрывались по присутствующим аллелям, определяющим пол, но не представляли собой близкого родства. Личинки Ephestia kuehniella Zeller (Lepidoptera: Pyralidae) служили хозяевами. Эксперименты и выращивание проводили при 25°C, 55% отн. вл. и 16:8 ч свет:темно.
Работали с 14-18-дневными девственными самками, ранее паразитировавшими на одном хозяине. Чтобы избежать вмешательства в выбор партнера по родству, отдельных самок одной популяции помещали с самцом из другой популяции и наоборот в пустую чашку Петри (ø 3,5 см). Самка могла либо принять ухаживающего самца, что приводило к совокуплению обычно в течение пяти минут («принятое» спаривание), либо отвергнуть его, согнув брюшко, ударив задними ногами или убежав при контакте. Если происходило отторжение, самку осторожно переносили в пластиковую пробирку объемом 250 мкл и помещали на лед на восемь минут, пока она не переставала двигаться. Неподвижную самку перенесли обратно в чашку Петри с тем же самцом, которого она сначала отвергла. Самец обычно подходил к ней немедленно, и совокупление происходило до того, как она снова восстанавливала полноценные движения («отвергнутое» спаривание; метод адаптирован из [25]). Чтобы проверить возможный эффект охлаждения, мы использовали контрольную группу самок, которых спаривали без охлаждения с одним из их собственных сыновей, которых они произвели девственницами. Эти самки были того же возраста, что и подопытные самки, и также имели один опыт откладывания яиц до спаривания. Поскольку гаплоидные сыновья наследуют один из половых аллелей своей матери, спаривания мать-сын по определению совпадают с точки зрения половых аллелей и, таким образом, должны приводить к тому, что примерно половина диплоидного потомства становится гомозиготной по локусу, определяющему пол (диплоидные самцы). Интересно, что около 1/2 самок, столкнувшихся с собственным сыном, без колебаний спаривались (и, таким образом, могли использоваться в качестве контроля эффектов охлаждения), несмотря на аллельное совпадение.
Неподвижную самку перенесли обратно в чашку Петри с тем же самцом, которого она сначала отвергла. Самец обычно подходил к ней немедленно, и совокупление происходило до того, как она снова восстанавливала полноценные движения («отвергнутое» спаривание; метод адаптирован из [25]). Чтобы проверить возможный эффект охлаждения, мы использовали контрольную группу самок, которых спаривали без охлаждения с одним из их собственных сыновей, которых они произвели девственницами. Эти самки были того же возраста, что и подопытные самки, и также имели один опыт откладывания яиц до спаривания. Поскольку гаплоидные сыновья наследуют один из половых аллелей своей матери, спаривания мать-сын по определению совпадают с точки зрения половых аллелей и, таким образом, должны приводить к тому, что примерно половина диплоидного потомства становится гомозиготной по локусу, определяющему пол (диплоидные самцы). Интересно, что около 1/2 самок, столкнувшихся с собственным сыном, без колебаний спаривались (и, таким образом, могли использоваться в качестве контроля эффектов охлаждения), несмотря на аллельное совпадение. Спаривание собственного сына не противоречит идее избежать аллельного соответствия, если мы рассмотрим идею о том, что принятие партнера меняется с опытом (например, [26, 27]): если сын является единственным потенциальным партнером, с которым сталкивается самка, это что вокруг нет сородичей и что шансы встретить неродственного самца очень малы.
Спаривание собственного сына не противоречит идее избежать аллельного соответствия, если мы рассмотрим идею о том, что принятие партнера меняется с опытом (например, [26, 27]): если сын является единственным потенциальным партнером, с которым сталкивается самка, это что вокруг нет сородичей и что шансы встретить неродственного самца очень малы.
После спаривания каждая самка паразитировала в общей сложности на пяти личинках-хозяевах в течение трех дней. Личинок-хозяев сразу же после извлечения самки исследовали для определения количества отложенных яиц. Через 10 дней ежедневно осматривали развивающееся потомство; Появляющихся ос определяли по полу и подсчитывали. Уровни плоидности потомства самцов определяли с помощью проточной цитометрии: отдельные головы ос гомогенизировали в буфере Гэлбрейта (21 мМ MgCl 2 , 30 мМ тринатрийцитрата дигидрата, 20 мМ MOPS, 0,1% Triton X-100, 1 мг/л РНКазы). 28]), фильтровали (40 мкм), окрашивали в течение 10 минут 15 мкл йодида пропидия (1,25 мг/мл) и загружали на проточный цитометр Coulter Epics XL-MCL (Beckman Coulter, Майами, Флорида, США). Мы использовали длину волны возбуждения 488 нм и полосовой фильтр 585 нм для обнаружения флуоресценции йодида пропидия. Образцы измеряли в закрытой области FSlog/FL2-log и FSlog/FL3-log до тех пор, пока не было достигнуто 2500–3000 отсчетов, используя Expo 32 ADC XL 4 Color (Beckman Coulter, США). Был применен порог для исключения очень мелкого мусора. Известные диплоиды (самки) и гаплоиды (самцы, произведенные матерями-девственницами) предоставили эталонные гистограммы (Рисунок 4), используемые для определения уровня плоидности произведенного потомства мужского пола. Самцы, для которых мы не смогли определить уровень плоидности (таблица 1), например. потому что либо не появлялось определенного гаплоидного пика, либо потому что отсутствие гаплоидного пика не могло быть подтверждено, использовались только в анализе выживаемости потомства.
Мы использовали длину волны возбуждения 488 нм и полосовой фильтр 585 нм для обнаружения флуоресценции йодида пропидия. Образцы измеряли в закрытой области FSlog/FL2-log и FSlog/FL3-log до тех пор, пока не было достигнуто 2500–3000 отсчетов, используя Expo 32 ADC XL 4 Color (Beckman Coulter, США). Был применен порог для исключения очень мелкого мусора. Известные диплоиды (самки) и гаплоиды (самцы, произведенные матерями-девственницами) предоставили эталонные гистограммы (Рисунок 4), используемые для определения уровня плоидности произведенного потомства мужского пола. Самцы, для которых мы не смогли определить уровень плоидности (таблица 1), например. потому что либо не появлялось определенного гаплоидного пика, либо потому что отсутствие гаплоидного пика не могло быть подтверждено, использовались только в анализе выживаемости потомства.
Эталонные гистограммы проточной цитометрии. Поскольку дупликация ДНК является обычным явлением для перепончатокрылых, гаплоидная особь (правая сторона) распознается по наличию гаплоидного пика в дополнение к диплоидному и тетраплоидному пикам. Диплоидная особь определяется отсутствием гаплоидного пика, в то же время присутствуют диплоидный и тетраплоидный пики (слева).
Диплоидная особь определяется отсутствием гаплоидного пика, в то же время присутствуют диплоидный и тетраплоидный пики (слева).
Полноразмерное изображение
Обобщенные линейные модели (GLM, [29]) были подобраны к данным, если не указано иное, с использованием статистических процедур в «R 2.15.2» [30] с пакетом «автомобиль» [31]. Для анализа доли оплодотворенных яиц, развивающихся как диплоидные самцы, мы использовали команду «cbind», чтобы учесть разное количество потомков, производимых каждой самкой. Распределение ошибок в этом тесте было квазибиномиальным с функцией логит-ссылки. Плодовитость самок и смертность потомства анализировали с помощью квазипуассоновских распределений ошибок и логарифмических функций связи.
Наши результаты показывают, что выбор самки может снизить вероятность соответствующего спаривания примерно до 1/4 того, что можно было ожидать при случайном спаривании в нашем эксперименте. Этот расчет основан на тех 18 парах, для которых мы смогли определить статус совпадения с их случайно назначенными партнерами (таблица 1): 0,4 из этих спариваний были совмещены. Среди самок, имевших «принятое» спаривание, совпадала лишь доля 0,1.
Среди самок, имевших «принятое» спаривание, совпадала лишь доля 0,1.
Сокращения
- CSD:
-
Дополнительное определение пола
- F1:
-
Первое поколение потомков
- GLM:
-
Обобщенная линейная модель
- МНС:
-
Главный комплекс гистосовместимости
- МОП:
-
3-(N-морфолино)пропансульфокислота
- П:
-
Родительское поколение
- СИ:
-
Самонесовместимость
Ссылки
-
Tregenza T, Wedell N: Генетическая совместимость, выбор партнера и модели отцовства: приглашенный обзор. Мол Экол. 2000, 9: 1013-1027.
Артикул КАС пабмед Google ученый
-
Mays HL, Hill GE: Выбор пары: хорошие гены против генов, которые хорошо подходят. Тенденции Экол Эвол. 2004, 19: 554-559.
Артикул пабмед Google ученый
-
Wegner KM, Kalbe M, Kurtz J, Reusch TBH, Milinski M: Отбор паразитов для иммуногенетической оптимальности. Наука. 2003, 301: 1343-1343.
Артикул КАС пабмед Google ученый
-
Ли Х.К., Симмонс Л.В., Родс Г.: Генетическое несходство, генетическое разнообразие и предпочтения в выборе партнера у людей. Эвол Бехав. 2010, 31: 48-58.
Артикул Google ученый
-
Kempenaers B: Выбор партнера и генетическое качество: обзор теории гетерозиготности. Ad Stud Behav. 2007, 37: 189-278.
Артикул Google ученый
-
Mays HL, Albrecht T, Liu M, Hill GE: Выбор самок для генетической комплементарности у птиц: обзор. Генетика. 2008, 134: 147-158.
Артикул пабмед Google ученый
-
Havlicek J, Roberts SC: MHC-коррелированный выбор партнера у людей: обзор. Психонейроэндокринология. 2009, 34: 497-512.
Артикул КАС пабмед Google ученый
-
Хант Дж., Бюссьер Л.Ф., Дженнионс М.Д., Брукс Р.: Что такое генетическое качество?. Тенденции Экол Эвол. 2004, 19: 329-333.
Артикул пабмед Google ученый
-
Нефф Б.Д., Питчер Т.Е. Генетическое качество и половой отбор: интегрированная структура для хороших генов и совместимых генов. Мол Экол. 2005, 14:19-38.
Артикул КАС пабмед Google ученый
-
Beye M, Hasselmann M, Fondrk MK, Page RE, Omholt SW: Ген csd является первичным сигналом полового развития медоносной пчелы и кодирует белок SR-типа. Клетка. 2003, 114: 419-429.
Артикул КАС пабмед Google ученый
-
Heimpel GE, de Boer JG: Определение пола у перепончатокрылых. Энн Рев Энтомол. 2008, 53: 209-230.
Артикул КАС Google ученый
-
Ван Вильгенбург Э., Дриссен Г., Бьюкебум Л.В.: Однолокусное определение комплементарного пола у перепончатокрылых: «неразумный» дизайн?. Фронт Зоол. 2006, 3: 1-10.1186/1742-9994-3-1.
Артикул ПабМед Центральный пабмед Google ученый
-
Speicher BR, Speicher KG: Наличие диплоидных самцов у Habrobracon brevicornis . Я Нат. 1940, 74: 379-382.
Артикул Google ученый
-
Bernatchez L, Landry C: Исследования MHC у немодельных позвоночных: что мы узнали о естественном отборе за 15 лет?. Дж. Эвол Биол. 2003, 16: 363-377.
Артикул КАС пабмед Google ученый
-
Hedrick PW, Gadau J, Page RE: Генетическое определение пола и исчезновение. Тенденции Экол Эвол. 2006, 21: 55-57.
Артикул пабмед Google ученый
-
Simmons LW: Феромональные сигналы для распознавания родства самками полевых сверчков, Gryllus bimaculatus .
Аним Бехав. 1990, 40: 192-195. Артикул Google ученый
-
Herzner G, Schmitt T, Heckel F, Schreier P, Strohm E: Братья пахнут одинаково: изменение полового феромона самцов европейских пчеловодов Philanthus triangulum F. (Hymenoptera: Crabronidae) и его значение для предотвращения инбридинга. Biol J Linn Soc. 2006, 89: 433-442.
Артикул Google ученый
-
Мецгер М., Бернштейн С., Хоффмайстер Т.С., Десуан Э. Ограничивает ли распознавание родственников и предотвращение спаривания сибса риск генетической несовместимости у осы-паразита? ПЛОС Один. 2010, 5: e13505-10.1371/journal.pone.0013505.
Артикул ПабМед Центральный пабмед Google ученый
-
Zayed A, Packer L: Комплементарная детерминация пола существенно повышает склонность к вымиранию гаплодиплоидных популяций.
Proc Nat Acad Sci USA. 2005, 102: 10742-10746. Артикул ПабМед Центральный КАС пабмед Google ученый
-
Хайн С., Поэтке Х.Дж., Дорн С.: Что останавливает «диплоидный мужской вихрь»? – имитационное исследование видов с одним локусом комплементарного определения пола. Экол мод. 2009 г., 220: 1663-1669.
Артикул Google ученый
-
Kitthawee S: Принудительно-контактное спаривание: метод экспериментов по скрещиванию с паразитоидом плодовой мухи, Diachasmimorpha longicaudata (Ashmead) (Hymenoptera: Braconidae). Биол Кон. 2008, 44: 73-78.
Артикул Google ученый
-
Bailey NW, Zuk M: Полевые сверчки меняют предпочтения в отношении спаривания, используя запомненную социальную информацию. Биол Лет. 2009 г., 5: 449-451.
Артикул Google ученый
-
Galbraith DW, Harkins KR, Maddox JM, Ayres NM, Sharma DP, Firoozabady E: Быстрый проточный цитометрический анализ клеточного цикла в интактных тканях растений. Наука. 1983, 220: 1049-1051.
Артикул КАС пабмед Google ученый
-
Nelder JA, Wedderburn RWM: Обобщенные линейные модели. JR Stat Soc A. 1972, 135: 370-384.
Артикул Google ученый
-
Основная группа разработчиков R: R: язык и среда для статистических вычислений. 2011, Вена, Австрия: R Foundation for Statistical Computing, 2140
Google ученый
 «>
«> Куропатка L: Выбор партнера увеличивает компонент приспособленности потомства плодовых мушек. Природа. 1980, 283: 290-291.
Артикул Google ученый
 «>
«> Wedekind C, Seebeck T, Bettens F, Paepke AJ: MHC-зависимые предпочтения в выборе партнера для людей. Proc R Soc London B. 1995, 260: 245-249.
Артикул КАС Google ученый
 «>
«> Бос Д.Х., Уильямс Р.Н., Гопуренко Д., Булут З., Девуди Дж.А. Выбор партнера в зависимости от состояния и репродуктивный недостаток самцов тигровых саламандр, расходящихся по MHC. Мол Экол. 2009, 18: 3307-3315.
Артикул пабмед Google ученый
 «>
«> Голдберг Э.Е., Кон Дж.Р., Ланде Р., Робертсон К.А., Смит С.А., Игик Б.: Селекция видов сохраняет самонесовместимость. Наука. 2010, 330: 493-495.
Артикул КАС пабмед Google ученый

 Аним Бехав. 1990, 40: 192-195.
Аним Бехав. 1990, 40: 192-195.  Proc Nat Acad Sci USA. 2005, 102: 10742-10746.
Proc Nat Acad Sci USA. 2005, 102: 10742-10746. 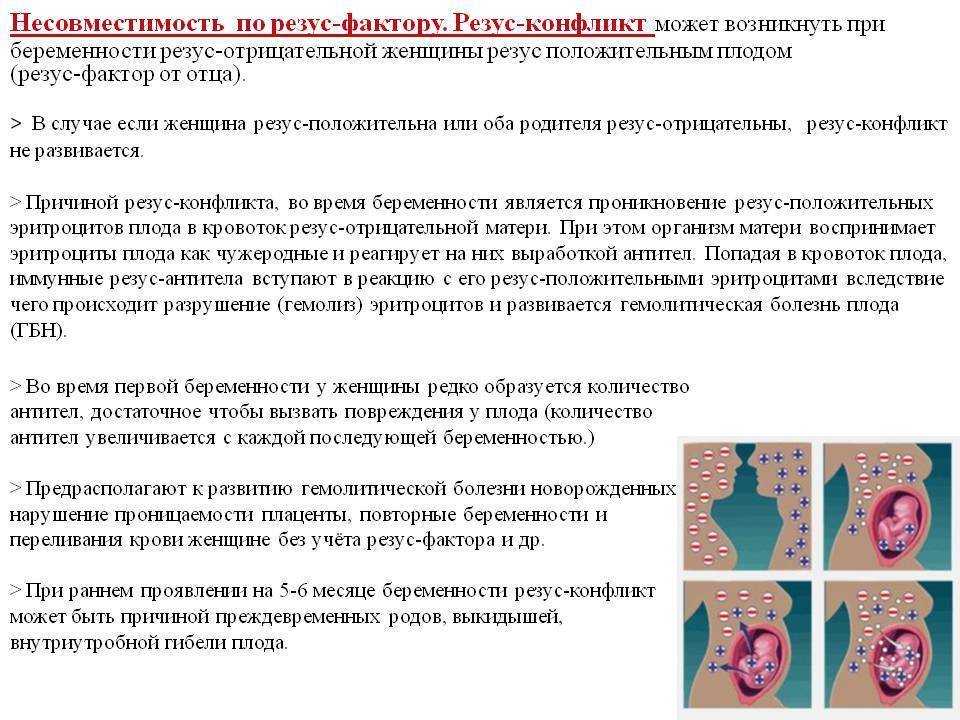 «>
«> Фаулер-Финн К.Д., Родригес Р.Л.: Опосредованная опытом пластичность в предпочтениях партнера: гарантия спаривания в изменчивой среде. Эволюция. 2012, 66: 459-468.
Артикул пабмед Google ученый
 «>
«> Фокс Дж., Вайсберг С.: (R) компаньон прикладной регрессии. 2011, Таузенд-Оукс, Калифорния: Sage, 2
Google ученый
Скачать ссылки
Благодарности
Исследование финансировалось ESF (BEPAR) с краткосрочным грантом на визит в AT, Федеральное министерство экономики и технологий Германии (BMWi) (KF 2152901MD8 для TSH) и Нидерландскую организацию для научных исследований (ALW 863.07.010 до JGdB). Мы благодарим B. Wührer и O. Zimmermann (AMW Nützlinge GmbH) за насекомых, W. van de Poll (морская биология, RUG) за помощь в проточной цитометрии и R. Amann, L.W. Бьюкебум, М. Дике и Б.Д. Ройтберга для обсуждения.
Информация об авторе
Авторы и организации
-
Популяционная и эволюционная экология, Институт экологии, Бременский университет, FB 2, Leobener Str., NW2, 28359, Бремен, Германия
Andra Thiel, Anne Comaeda S Hoffmeister
-
Эволюционная генетика, Центр экологических и эволюционных исследований, Гронингенский университет, почтовый ящик 11103, 9700, CC Гронинген, Нидерланды
Jetske G de Boer
-
Лаборатория энтомологии, Вагенингенский университет, P.
O. Box 8031, 6700, EH Wageningen, Нидерланды Jetske G de Boer
 O. Box 8031, 6700, EH Wageningen, Нидерланды
O. Box 8031, 6700, EH Wageningen, Нидерланды Авторы
- Andra Thiel
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Scholar
- Anne C Weeda
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Академия
- Jetske G de Boer
Посмотреть публикации автора
Вы также можете искать этого автора в PubMed Google Scholar
- Thomas S Hoffmeister
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Scholar
Автор, ответственный за переписку
Андра Тиль.
Дополнительная информация
Конкурирующие интересы
Авторы заявляют, что у них нет конкурирующих интересов.
Вклад авторов
AT инициировал и руководил проектом, ACW проводил эксперименты, JGdB руководил проточной цитометрией. AT и TSH выполнили статистику. AT написал первый черновик, и все авторы внесли свой вклад в интерпретацию результатов и улучшение статьи. Все авторы прочитали и одобрили окончательный вариант рукописи.
AT и TSH выполнили статистику. AT написал первый черновик, и все авторы внесли свой вклад в интерпретацию результатов и улучшение статьи. Все авторы прочитали и одобрили окончательный вариант рукописи.
Оригинальные файлы изображений, представленные авторами
Ниже приведены ссылки на оригинальные файлы изображений, представленные авторами.
Оригинальный файл авторов для рисунка 1
Оригинальный файл авторов для рисунка 2
Оригинальный файл авторов на рисунок 3
Оригинальный файл авторов
0 Rights
.
Эта статья опубликована по лицензии BioMed Central Ltd. Это статья с открытым доступом, распространяемая в соответствии с условиями лицензии Creative Commons Attribution License (http://creativecommons.org/licenses/by/2.0), которая разрешает неограниченное использование, распространение , а также воспроизведение на любом носителе при условии правильного цитирования оригинальной работы.