
Конъюнктивный моллюск: Контагиозный моллюск у детей — причины, симптомы, диагностика и лечение контагиозного моллюска у ребенка в Москве в клинике «СМ-Доктор»
Контагиозный моллюск у детей — причины, симптомы, диагностика и лечение контагиозного моллюска у ребенка в Москве в клинике «СМ-Доктор»
ПОЛУЧИТЬ КОНСУЛЬТАЦИЮ Содержание:Общие сведения
Симптомы у детей
Причины
Диагностика
Лечение
Профилактика
Контагиозный моллюск — вирусная инфекция с характерными кожными проявлениями. Чаще болеют дети до 10 лет и взрослые от 20 до 30 лет. Лечением заболевания занимаются педиатры, дерматологи.
Общие сведения
Контагиозный моллюск у детей как патология отмечен в международной классификации болезней (МКБ-10) под кодом В08.1. Хроническая форма болезни у подростков является поводом для отсрочки или иногда недопущения к службе в армии.При напряженном иммунитете заболевание проходит самостоятельно через 3-6 месяцев. Если иммунная система слабая, при отсутствии лечения патология переходит в хроническую форму.
Лечение контагиозного моллюска у детей, если начато вовремя, в основном завершается успешно. Прогноз благоприятный, осложнения и рецидивы случаются редко. Справиться с заболеванием легко – важно укрепить детский иммунитет.
Симптомы контагиозного моллюска
После попадания вируса в организм может пройти от 2 недель до полугода, прежде чем заболевание проявит себя. Для патологии характерны высыпания, внешне похожие на обычные папулы (прыщики) телесного цвета. Уплотнения напоминают узелки или подкожные шарики. По оттенку они почти не отличаются от основного цвета кожного покрова, хотя могут быть чуть белее. Верхушка жемчужного цвета. По мере развития узелка в центре появляется вдавление, напоминающее по виду «пупок».Прыщики бывают одиночные или сливаются в группы. Они располагаются в основном на лице, на коже верхней части туловища. Реже встречаются на ногах, бедрах или ягодицах. Если раздавить пузырек, из него выделится сухая творожистая масса.
Нельзя определенно сказать, чем опасен контагиозный моллюск у ребенка.
Причины контагиозного моллюска
Контагиозный моллюск вызывается одноименным вирусом. Выделяют 4 типа, но патологию чаще провоцируют 1-й и 2-й типы. Микроорганизм относится к группе вирусов оспы и опасен только для человека: животные к нему нечувствительны.Высыпания на лице и теле у детей вызывает обычно вирус 1-го типа. Заражение вторым типом, как правило, проявляется в виде характерных высыпаний в зоне половых органов. Для обоих микроорганизмов характерен длительный инкубационный период, доходящий до полугода.
Передача вирусов у детей происходит контактным путем. Если в коллективе есть больной ребенок, у других малышей высока вероятность подхватить вирус. Заражение реализуется такими путями:
- через средства личной гигиены;
- при купании в общем бассейне, водоеме;
- при вдыхании пыли (вирус хорошо сохраняется в земле, пыли).
Повышают риск заражения ослабленный иммунитет, несоблюдение гигиены, аллергическая реакция, лечение иммунодепрессантами, дерматологические болезни. Особенно опасен моллюск для детей с положительным ВИЧ-статусом.
Диагностика контагиозного моллюска
Заболевание сложно выявить самостоятельно, поскольку узелки на коже напоминают обычные прыщики. Если высыпания единичные, взрослые чаще всего не обращают внимания на эти признаки и полагают, что они пройдут сами.В клинике «СМ-Доктор» работают только опытные врачи, которым не составит труда поставить правильный диагноз еще на первом приеме.
Для подтверждения диагноза берут соскоб кожи и изучают материал под микроскопом. При заражении вирусом в цитоплазме клеток заметны моллюсковые тельца.
Диагноз «контагиозный моллюск» еще не означает, что у ребенка есть серьезные проблемы со здоровьем.
Лечение контагиозного моллюска
Если у взрослых с сильным иммунитетом болезнь можно не лечить, то дети нуждаются в курсе терапии. После консультации педиатра ребенок должен посетить дерматолога.Чем лечить контагиозный моллюск у детей, зависит от выбора врача:
- Можно удалять содержимое «прыщей» пинцетом и острой ложечкой. После процедуры пораженные зоны смазывают йодом.
- Можно прижигать лазером или током. Такой метод удаления возбудителя называется диатермокоагуляция.
- Можно прижигать высыпания сухим льдом или жидким азотом.
Если сыпь занимает значительные участки тела, показана системная терапия –иммуностимулирующие медикаменты и антибиотики. Детям до 10 лет прописывают только иммуномодуляторы, поскольку антибиотики тетрациклинового ряда им противопоказаны.
Важно! Нельзя лечить моллюск народными средствами: травами, прижиганиями. Подобные меры только увеличивают зону поражения на коже. Лучше обратиться к врачу и получить традиционное лечение с доказанной эффективностью.
Профилактика контагиозного моллюска
Избежать заражения вирусом сложно. Это может случиться в любой момент, поскольку больной человек может оказаться рядом в транспорте, встретиться при посещении больницы и других общественных учреждений, при пользовании бассейном и т.д. Но крепкая иммунная система предотвратит развитие заболевания. Оградить ребенка от вируса можно соблюдением мерами профилактики:- пополнять рацион малыша фруктами и овощами, придерживаться правильного питания;
- принимать витаминные комплексы;
- чаще проветривать помещение, гулять на свежем воздухе;
- заниматься физическими упражнениями;
- избегать мест большого скопления людей;
- следить за здоровьем ребенка и вовремя лечить вирусные болезни;
- соблюдать правила гигиены;
- пользоваться индивидуальными средствами по уходу за своим телом.
Контагиозный моллюск у детей не опасен, его проявления легко излечиваются. Но чтобы избежать рецидивов в будущем, нужно следить за иммунитетом, не переохлаждаться, полноценно питаться и вести здоровый образ жизни. В клинике «СМ-Доктор» опытные специалисты быстро и достоверно установят диагноз, пропишут самые действенные схемы лечения и порекомендуют меры профилактики рецидивов.
Врачи:
Бозунов Алексей Викторович
Детский дерматолог, детский миколог
Зуева Ксения Михайловна
Смолева Мария Борисовна
Детский дерматолог высшей категории, детский миколог, детский трихолог, детский косметолог
Королькова (Симонович) Полина АскольдовнаДетский дерматолог, детский трихолог, детский миколог
Чекрыгина Марина Вячеславовна
Детский дерматолог, детский миколог, врач высшей категории. Заместитель главного врача по медицинской части в детском отделении на Волгоградском проспекте
Заместитель главного врача по медицинской части в детском отделении на Волгоградском проспекте
Кулешов Андрей Николаевич
Детский дерматолог, детский миколог, детский трихолог
Цуканов Сергей Владимирович
Детский дерматолог, детский миколог, детский трихолог
Багрянцева Мария Евгеньевна
Детский дерматолог, детский миколог, детский косметолог, детский трихолог, врач высшей категории
Нехорошева Инна Андреевна
Детский дерматолог, детский миколог, детский трихолог
Записаться на прием Детская клиника м. ВДНХ
ВДНХ
Симакова (Мухина) Екатерина Сергеевна
Детский дерматолог, детский миколог, детский косметолог, к.м.н.
Уварова Елена Анатольевна
Детский дерматолог, детский трихолог
Антонова Ольга Александровна
Детский дерматолог, детский миколог, детский трихолог
Стражников Руслан Андреевич
Детский дерматолог, детский миколог, детский трихолог, детский косметолог, врач высшей категории
Быханова Ольга Николаевна
Детский дерматолог, детский миколог
Записаться на прием Детская клиника в г. Солнечногорск, ул. Красная
Солнечногорск, ул. Красная
Жукова Кристина Игоревна
Детский дерматолог
Записаться на приемЗаписаться на прием
Мы гарантируем неразглашение персональных данных и отсутствие рекламных рассылок по указанному вами телефону. Ваши данные необходимы для обеспечения обратной связи и организации записи к специалисту клиники.
Контагиозный моллюск. Откуда на коже появляются странные пузырьки | Здоровая жизнь | Здоровье
Не только на дне морском
Слово «контагиозный» значит «заразный, передающийся при прикосновении». Это полностью объясняет, почему возникает болезнь. Дети обычно заражаются контагиозным моллюском в яслях и детском саду, на детской площадке и в школе, соприкасаясь в игре с больным ребёнком, а также через общие игрушки и книги. Можно заразиться, пользуясь чужим полотенцем, губкой, мочалкой, расчёской, постельным бельём. В летних лагерях это происходит, если дети, желая пофорсить, берут чужую одежду.
Взрослые могут подхватить моллюска в бассейне, бане, фитнес-центре. Но самый распространённый путь заражения – сексуальный контакт. Не случайно у заболевания два пика: у детей больше всего случаев приходится на возраст от трёх до девяти лет, у подростков и взрослых – на 17–24 года, то есть на начало половой жизни. Интересно, что пик заражённости у взрослых в Европе пришёлся на 60–80‑е годы прошлого столетия, когда до «старушки» докатились отголоски американской «сексуальной революции».
Интересно, что пик заражённости у взрослых в Европе пришёлся на 60–80‑е годы прошлого столетия, когда до «старушки» докатились отголоски американской «сексуальной революции».
Иногда контагиозным моллюском болеют люди среднего возраста и пожилые. Скорее всего, они заразились ещё в детстве или в молодости, но заболевание проявилось, лишь когда ослаб иммунитет.
Не простой прыщик
Внешне контагиозный моллюск выглядит как прыщик. Пожалуй, нет части тела, кроме ладоней и подошв, где бы он ни обитал. У детей это руки, лицо и шея. У подростков и взрослых – наружные половые органы, лобок, промежность, внутренняя поверхность бёдер, низ живота.
Чтобы поставить диагноз «контагиозный моллюск», опытному врачу не нужно проводить сложных исследований. Достаточно внимательно рассмотреть объект жалоб – высыпания на коже. Они бывают телесного, розоватого или желтоватого цвета размером от полутора до десяти миллиметров (хотя иногда размер может быть и несколько сантиметров). Встречается и так называемая милиарная форма – множество крошечных высыпаний.
У пузырьков перламутровая верхушка, на которой, если рассмотреть под лупой, виден вдавленный центр. При надавливании пинцетом (именно так происходит диагностика) из центра узелка покажется содержимое – кашицеобразная масса, которая содержит округлые тельца, похожие на маленьких моллюсков.
«Осьминогов» под кожей нет
Именно из-за внешнего сходства содержимого узелков болезнь получила такое название. На самом деле никакие моллюски под кожу не забираются, а изменения на ней вызваны действием вируса Molitor hominis. Известно два его типа. На долю «детского» типа 1-го приходится до 96,6% всех случаев заболевания. У взрослых виновником заражения является передаваемый при интимных контактах 2-й тип вируса, на его совести – 3,4% случаев заболевания. Заразившись, человек навсегда становится носителем вируса моллюска, но для окружающих он не опасен. Заразным он становится, только если вирус проснётся и появятся высыпания.
От заражения до появления моллюсков на коже проходит от двух недель до нескольких месяцев.
Вирус находится в содержимом пузырьков, поэтому их нельзя ковырять, чесать, сбривать или пытаться ещё как-то удалять самостоятельно. У большинства людей контагиозный моллюск исчезает без всякого лечения максимум за полгода. Но далеко не все согласны ждать так долго, особенно если моллюски облюбовали лицо или шею, и предпочитают удалить их у врача дерматолога-косметолога.
Народные целители советуют смазывать моллюск соком чистотела и одуванчика, накладывать на них кашицу из чеснока. Эффективность таких средств не доказана, ведь лечиться придётся долго – примерно столько же, сколько нужно, чтобы моллюск исчез самостоятельно. Есть и другая причина, по которой не стоит уповать на народную медицину. Моллюск можно спутать с другими заболеваниями, в том числе с опухолями кожи, которые требуют совершенно другого лечения.Лазер против пинцета и мази
Самый традиционный способ изгнать моллюска с кожи – физическое устранение его с помощью пинцета или выскабливания с помощью так называемой ложечки Фолькмана. Затем места обитания моллюска смазывают йодом, зелёнкой, насыщенным раствором марганцовки. После этого моллюски какое-то время продолжают появляться, и их надо вновь и вновь удалять и прижигать. Чтобы избежать рецидивов, надо обрабатывать места высыпания противовирусными мазями.
Взрослые обычно стойко проходят через такое удаление, а вот дети бурно протестуют, потому что больно. При этом все попытки сделать удаление контагиозного моллюска менее болезненным пока неудачны.
Этот же недостаток есть и у таких способов борьбы с высыпаниями, как электро- и диатермокоагуляция (прижигание токами высокой частоты) и криодеструкция (вымораживание при помощи жидкого азота или охлаждённого до температуры минус 150–190 градусов воздуха).
Какое-то время врачам казалось, что хорошее решение – удалять моллюска медикаментозными методами. Для этого поражённые места необходимо смазывать препаратами нитрата серебра, интерфероном, прижигающими растворами. Но популярным этот метод не стал: он требует от пациента аккуратности и настойчивости, а ждать результата приходится по месяцу и больше.
Для этого поражённые места необходимо смазывать препаратами нитрата серебра, интерфероном, прижигающими растворами. Но популярным этот метод не стал: он требует от пациента аккуратности и настойчивости, а ждать результата приходится по месяцу и больше.
Гораздо эффективнее удаление контагиозного моллюска с помощью хирургического лазера. Его преимущество прежде всего в том, что удаляются только больные ткани, здоровые остаются неповреждёнными. Через 3–4 дня корочки, образовавшиеся на месте удаления, отпадают, никаких следов не остаётся. Это особенно важно, если моллюска пришлось изгонять с лица или шеи. На время лечения больного ребёнка нельзя водить в сад или школу, взрослым запрещены сексуальные контакты, половые партнёры должны лечиться вместе. Членам семьи и тем, кто близко контактировал с заболевшим, стоит сделать профилактику интерфероном.
Смотрите также:
Офтальмология – центр офтальмологии в Санкт-Петербурге
directions
На ухудшение зрения могут повлиять физическое напряжение глазных мышц и нервов, длительная работа за компьютером, недостаточность освещения или световое напряжение, стрессы, психическое и умственное напряжение, травмы шеи и головы, неправильная работа кишечника.
Функции глаз могут страдать также из-за перенесенных ранее неврологических заболеваний (атрофия зрительных нервов), сахарного диабета (рецидивирующий гемофтальм, диабетическая ретинопатия), атеросклероза, почечной патологии, патологии беременности, гипертонической болезни, заболеваний крови.
Врачи-специалисты
Врач-офтальмолог
Врач-офтальмолог
Врач-офтальмолог
Врач-офтальмолог
В настоящее время на сайте ведутся работы по изменению прайс-листа, актуальную информацию уточняйте по тел: 640-55-25 или оставьте заявку, с Вами свяжется оператор.
Цены на услуги
- Ретиноскопия 100a
- Исследование остроты зрения 270a
- Исследование цветоощущения 270a
- Гониоскопия 300a
- Визометрия 400a
- Офтальмотонометрия (2 глаза) ВГД по Маклакову 400a
- Подбор очковой коррекции зрения 400a
- Рефрактометрия 400a
- Офтальмодинамометрия 400a
- Получение мазка содержимого конъюнктивальной полости и слезоотводящих путей 100a
- Инстилляция капель, закладывание мази 200a
- Соскоб конъюнктивы 250a
- Промывание конъюнктивной полости 400a
- Промывание слезных путей 660a
- Удаление инородного тела с роговицы 2320a
- Прием (осмотр, консультация) врача — офтальмолога повторный 1100a
- Прием (осмотр, консультация) врача — офтальмолога первичный 1200a
- Прием (осмотр, консультация) врача — офтальмолога первичный (Визометрия, Исследование глазного дна, Биомикроскопия переднего отрезка глаза, Офтальмотонометрия ВГД по Маклакову) 1500a
Информация и цены, представленные на сайте, являются справочными и не являются публичной офертой.
Наши клиники в Санкт-Петербурге
Медицентр Юго-ЗападПр.Маршала Жукова 28к2
Кировский район
- Автово
- Проспект Ветеранов
- Ленинский проспект
Получить подробную информацию и записаться на прием Вы можете по телефону +7 (812) 640-55-25
Поводом обратиться к офтальмологу могут стать:
- Ухудшение зрения;
- Боли в глазных яблоках;
- Сухость склер или постоянное слезотечение;
- Головные боли.
Квалифицированные врачи-офтальмологи медицинского центра «Медицентр» проведут диагностику зрения, составят индивидуальные профилактические программы, окажут экстренную помощь при лёгких травмах органов зрения, а также при попадании в глаза инородных тел.
В нашем медицинском центре проводятся:
- Осмотр и диагностика состояния глазного яблока;
- Определение остроты зрения;
- Измерение внутриглазного давления;
- Подбор очков.
Рекомендуется проходить консультацию у врача-офтальмолога раз в год людям любого возраста в профилактических целях, особое внимание здоровью глаз следует уделять пациентам старше 40 лет.
На консультацию к офтальмологу желательно принести с собой данные предыдущих осмотров окулиста и других специалистов.
871,1307,948,832,1328,1300
Смерткин Алексей Сергеевич 11.02.2021 19:40medi-center.ru
Хочу выразить благодарность врачу Саранчину Александру за качественный осмотр и рекомендации в лечении. Успехов Вам, Александр и профессионально развития.
Добрый день !
Хочу всех поздравить с наступившим Новым годом, пожелать всем хочу здоровья и благополучия.Но особенно хочу поздравить доктора клиники в Мурино , на Охтинской аллее-это доктор Султанга Валерия Дмитриевна.
Уважаемая Валерия Дмитриевна!
Хочу выразить Вам свою огромную благодарность за колоссальную помощь, за отзывчивость, за качественное лечение, за врачебную этику и за профессионализм своего дела.В нелегкий для моей жизни момент мне очень повезло встретить мудрого и чуткого специалиста. Спасибо Вам огромное за поддержку и профессиональное лечение.
Спасибо Вам за Вашу бесконечную доброту и теплое отношение. Желаю Вам всего самого наилучшего и только благодарных пациентов.От всей души хочу сказать Вам «большое спасибо». Вы — врач от Бога.Благодарю Вас за Ваше доброе сердце , за оказанную помощь, за возможность полноценной жизни и за добрую надежду моей души. Желаю Вам достатка и здоровья в Новом году , сил и безграничных возможностей, подаренных судьбой.
Очень хотелось бы , что бы руководство клиники замечала таких докторов и каким то образом премировала таких уникальных по своей доброте людей , ведь такие доктора как Валерия Дмитриевна-уникальны , несмотря на то что это молодой доктор- она к каждому пациенту относится как к родному, со своим подходом, и восхищает своей самоотдачей любимому делу!
Еще раз благодарю доктора и от всей души ей всех благ и сил и терпения. С уважением Анна Н.
С уважением Анна Н.
medi-center.ru
Огромная благодарность неврологу Соловьёву Даниилу Петровичу! Настоящий доктор, побольше бы таких докторов!
Груздев Алексей Анатольевич 09.08.2020 16:34medi-center.ru
Прошел эндоскопическое обследование у группы врачей в составе: Мордвинцевой Анны Ахатовны, Суйдюмова Тимура Руслановича, Локотковой Александры Владимировны. Отношение к пациенту внимательное, ответственное. Работу выполняют профессионально, уверенно, без суеты. Спасибо!
Добры день всем, хочу выразить благодарность врачу-неврологу Братановой Ирине Валерьевне. На прием пришел с параличом лицевого нерва, левая сторона лица была практически парализована. Как оказалось, иглорефлексотерапия вполне успешно лечит такие болезни, спустя пару сеансов были заметны изменения в лучшую сторону, а на 8-9-м все пришло в норму, считаю это не только своей заслугой, но и врача :). Цены на ИРТ по району кстати довольно умеренные, а клиника на ул.Поликарпова 6 произвела хорошее впечатление.
Хочу выразить большую благодарность клинике на аллее Поликарпова 6к2, всегда вежливые, доброжелательные девушки на ресепшн. Помогли удобно состыковать время приёма врачей. А также отдельно поблагодарить замечательнейшего доктора Гиндрюк Василия Васильевича за профессионализм, заботу и внимание! Наблюдаться у него одно удовольствие!!!
Отделение офтальмологии — диагностика и лечение нарушений зрения у взрослых и детей, коррекция с помощью очков и контактных линз в Клиническом госпитале на Яузе |Москва
Во время диагностического обследования применяется современное оборудование. Мы используем стерильные одноразовые расходные материалы, что обеспечивает полную безопасность пациента.
Мы используем стерильные одноразовые расходные материалы, что обеспечивает полную безопасность пациента.
Цифровое изображение сетчатки
Немидриатическая фундус-камера — прибор для обследования и фотографирования сетчатки, кровеносных сосудов глазного дна и зрительного нерва. Цифровое изображение сетчатки — важный инструмент мониторинга здоровья глаз у людей любого возраста, поскольку обеспечивает наиболее точную визуализацию исследуемых структур.
ПодробнееТонометрия
Бесконтактный пневмотонометр Reichert 7 CR автоматическое, очень точное измерение внутриглазного давления с учетом биомеханических свойств роговицы. Используется воздушный поток. Метод не требует использования капель для анестезии, подходит для пациентов после рефракционных хирургических вмешательств.
Тонометр Icare — для измерения внутриглазного давления без применения воздушного потока. Используется как у взрослых, так и у детей любого возраста. Анестезия не нужна. Прибор обеспечивает получение высокоточных результатов и комфорт пациента.
ПодробнееЦифровое изображение передней поверхности глаза
Получается при помощи цифровой щелевой лампы SL C.S.O., оснащённой фото -/ видеорегистратором. Может быть записано и сохранено в цифровом качестве с самым высоким уровнем детализации роговицы, радужной оболочки, склеры и век. Используется для последующего мониторинга таких заболеваний, как катаракта, дистрофия роговицы, кератоконус, иридоциклит, птеригиум, невусы или пигментные образования, синдром сухого глаза, а также для подбора специальных контактных линз для коррекции сложного зрения (астигматизма, кератоконуса), после операций на роговице и при подборе ортокератологических «ночных» контактных линз для коррекции миопии.
ПодробнееСканирующая топография роговицы
Корнеотопограф Sirius позволяет рассчитывать наиболее точные карты передней и задней поверхности роговицы благодаря использованию двух методов измерения в одном приборе. На основании полученных данных строится полная пахиметрическая карта для определения толщины роговицы и исключения противопоказаний перед рефракционными операциями, а также в экспертной диагностике кератоконуса и оценке его прогрессирования.
На основании полученных данных строится полная пахиметрическая карта для определения толщины роговицы и исключения противопоказаний перед рефракционными операциями, а также в экспертной диагностике кератоконуса и оценке его прогрессирования.
Неотъемлемый этап подбора контактных линз при астигматизме, после операций на роговице, при подборе ортокератологических «ночных» контактных линз. Точное определение глубины и формы угла передней камеры необходимо при диагностике глаукомы.
Проверка зрения и определение рефракции
Измерение рефракции (оптической силы роговицы и хрусталика) — это базовое обследование при проверке зрения у пациента любого возраста.
Подробнее| Наименование | Цена |
| Тест Ширмера | 187,00р. |
| Периметрия статическая | 156,00р. |
| Исследование бинокулярного зрения | 229,00р. |
| Офтальмоскопия | 437,00р. |
| Исследование аккомодации | 161,00р. |
| Экзофтальмометрия | 177,00р. |
| Измерение угла косоглазия | 187,00р. |
| Биомикрофотография глазного дна с использованием фундус-камеры | 312,00р. |
| Диафаноскопия глаза | 239,00р. |
| Гониоскопия | 385,00р. |
| Введение кортикостероидов в халязион | 270,00р. |
| Офтальмохромоскопия | 312,00р. |
| Биомикроскопия глаза | 135,00р. |
| Флюоресцеиновая инстилляционная проба | 104,00р. |
| Скарификация и туширование роговичных очагов воспаления | 146,00р. |
| Скиаскопия | 229,00р. |
| Забор материала для исследования на демодекс | 260,00р. |
| Зондирование слезных канальцев, активация слезных точек | 229,00р.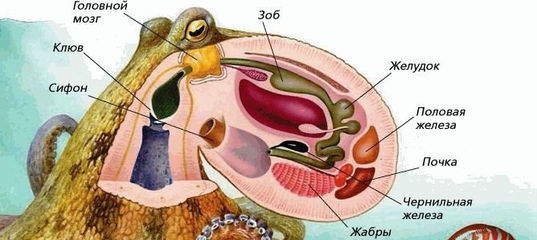 |
| Канальцевая проба (носовая проба, слезно-носовая проба) | 229,00р. |
| Определение силы оптических линз на диоптриметре | 94,00р. |
| Подбор сложной очковой коррекции зрения | 520,00р. |
| Зондирование слезно-носового канала | 364,00р. |
| Массаж века (один глаз) | 229,00р. |
| Субконъюнктивальная инъекция | 187,00р. |
| Пара- и ретробульбарные инъекции | 239,00р. |
| Промывание слезных путей | 322,00р. |
| Инстилляция лекарственных веществ | 62,00р. |
| Удаление инородного тела роговицы | 374,00р. |
| Эпиляция ресниц | 291,00р. |
| Удаление контагиозного моллюска, вскрытие малых ретенционных кист век и конъюнктивы, ячменя, абсцесса века | 374,00р. |
| Промывание конъюнктивной полости | 166,00р. |
| Исследование цветоощущения | 57,00р. |
| Локализация разрывов, инородных тел сетчатки | 333,00р. |
| Офтальмотонометрия | 146,00р. |
| Нагрузочно-разгрузовные пробы для исследования регуляции внутриглазного давления | 229,00р. |
| Исследование конвергенции | 94,00р. |
| Компьютерная периметрия | 1 092,00р. |
| Кампиметрия | 172,00р. |
| Рефрактометрия | 385,00р. |
| Авторефрактометрия с узким зрачком | 156,00р. |
| Офтальмометрия | 229,00р. |
| Тонометрия глаза бесконтактная | 146,00р. |
| Никтометрия (определение сумеречного зрения) | 146,00р. |
| Обследование на проекторе испытательных объектов | 229,00р. |
| Исследование диплопии | 572,00р. |
| Снятие кожных швов | 156,00р. |
| Снятие коньюнктивальных швов | 572,00р. |
| Снятие роговичных швов | 1 258,00р. |
| Послеоперационная обработка глаза | 229,00р. |
| Подбор очковой коррекции зрения | 333,00р. |
| Ультразвуковое исследование глазного яблока | 770,00р. |
| Биомикроскопия глазного дна | 302,00р. |
| Удаление инородного тела конъюнктивы | 364,00р. |
| Офтальмоскопия с широким зрачком | 426,00р. |
| Определение сумеречного зрения (мезотест) | 156,00р. |
| Периметрия тотальная | 676,00р. |
| Периметрия центральная | 624,00р. |
| Периметрия на цвета | 780,00р. |
| Проведение гониоскопической компрессионной пробы Форбса | 780,00р. |
| Инстилляция лекарственных веществ в конъюнктивную полость | 416,00р. |
| Удаление контагиозных моллюсков | 1 664,00р. |
| Периметрия периферическая | 624,00р. |
| Удаление инородного тела век | 536,00р. |
| Определение ретинальной остроты зрения | 146,00р. |
| Получение мазка содержимого конъюнктивальной полости и слезоотводящих путей | 676,00р. |
| Определение характера зрения, гетерофории | 130,00р. |
| Эластотонометрия | 172,00р. |
| Осмотр периферии глазного дна с использованием трехзеркальной линзы Гольдмана | 416,00р. |
| Удаление инородного тела из переднего сегмента глаза | 510,00р. |
| Промывание слезоотводящих путей | 291,00р. |
| Удаление контагиозного моллюска век и периорбитальной области | 468,00р. |
| Компьютерная рефрактометрия | 260,00р. |
| Наложение повязки при операциях на органе зрения | 343,00р. |
| Исследование переднего сегмента глаза методом бокового освещения | 555,00р. |
| Исследование сред глаза в проходящем свете | 555,00р. |
| Визометрия | 312,00р. |
| Определение рефракции с помощью набора пробных линз | 441,00р. |
| Определение дефектов поверхности роговицы | 555,00р. |
Страница не найдена — ГОБУЗ «МГП № 2»
17 МАРТ 2021 / СРЕДА
Компенсация стоимости путёвок медработникам и водителям СМП
17 МАРТ 2021 / СРЕДА
17 МАРТ 2021 / СРЕДА
17 МАРТ 2021 / СРЕДА
17 МАРТ 2021 / СРЕДА
17 МАРТ 2021 / СРЕДА
17 МАРТ 2021 / СРЕДА
17 МАРТ 2021 / СРЕДА
17 МАРТ 2021 / СРЕДА
17 МАРТ 2021 / СРЕДА
17 МАРТ 2021 / СРЕДА
17 МАРТ 2021 / СРЕДА
11 ДЕКАБРЬ 2019 / СРЕДА
Доступная среда
03 ДЕКАБРЬ 2019 / ВТОРНИК
Памятка о гарантиях оказания бесплатной медицинской помощи
17 ДЕКАБРЬ 2019 / ВТОРНИК
Частые вопросы и ответы
17 МАРТ 2021 / СРЕДА
17 МАРТ 2021 / СРЕДА
17 МАРТ 2021 / СРЕДА
17 МАРТ 2021 / СРЕДА
17 МАРТ 2021 / СРЕДА
17 МАРТ 2021 / СРЕДА
ФГБУ ‘Поликлиника №4’ Управление делами Президента РФ
Уважаемые посетители!
Поликлиника является уникальным лечебно-профилактическим учреждением, по существу семейным,
поскольку в стенах его медицинскую помощь одновременно получают прикрепленные к поликлинике
взрослые и дети.
Поликлиника входит в состав Управления делами Президента Российской Федерации и осуществляет свою деятельность, используя опыт и лучшие традиции Кремлевской медицины.
Высокий уровень оказания профилактической и лечебно-диагностической помощи, современная материально-техническая база, высокий уровень врачей и средних медицинских работников, доброжелательное отношение к своим пациентам и проблемам их здоровья создали поликлинике хорошую репутацию и заслуженный авторитет не только у пациентов, но и у страховых медицинских организаций и медицинской общественности. Поликлиника постоянно динамически развивается и обновляется в соответствии с требованиями времени и современных научных достижений.
В соответствии с имеющейся лицензией, поликлиника:
- На многие виды медицинской деятельности
- Оформляет медицинские справки и заключения:
- на право управления автотранспортом и ношения оружия;
- по результатам профилактических осмотров декретированных контингентов и периодических осмотров лиц с вредными и опасными для жизни условиями труда;
- об отсутствии противопоказаний для посещения бассейна;
- санаторно-курортные карты;
- Оказывает косметологические услуги;
- Проводит диспансеризацию, оформление паспорта здоровья, консультации пациентов на дому.
В составе взрослого корпуса имеются отделения терапевтического и хирургического профилей,
женская консультация, дневной стационар, стоматологическое отделение; диагностическая служба
в виде отделения лучевой диагностики, кабинета функциональной диагностики, эндоскопического
отделения, а так же клинико-диагностической лаборатории, в которой выполняются практически
все виды исследований в короткие сроки. Отделение медицинской реабилитации и спортивной
медицины представлено различным спектром оказываемых услуг (физиотерапия, бальнеотерапия,
иглорефлексотерапия, массаж, остеопатия, ЛФК и др.)
Отделение медицинской реабилитации и спортивной
медицины представлено различным спектром оказываемых услуг (физиотерапия, бальнеотерапия,
иглорефлексотерапия, массаж, остеопатия, ЛФК и др.)
В состав Детского корпуса входят: педиатрическая служба, отделение специализированной медицинской помощи, отделение медицинской реабилитации, восстановительной и спортивной медицины и лучевой диагностики, а также стоматология.
В Педиатрической службе нашим маленьким пациентам предлагаются в том числе:
- «Первый месяц жизни»
- «Наблюдение детей до года в поликлинике и на дому»
- «Идем в детский сад»
- «Идем в школу»
- «Поступаем в ВУЗ»
- «Здоровое поколение»
- «Хочу говорить!»
Отделение специализированной педиатрической медицинской помощи оказывает комплексную медицинскую помощь с привлечением узкопрофильных специалистов высокого уровня детям от рождения до 18 лет.
Основным уставным видом деятельности ФГБУ «Поликлиника № 4» является оказание медицинской помощи определенному контингенту — лицам, медицинское обслуживание которых в соответствии с законодательством возложено на Управление делами Президента Российской Федерации, в том числе в рамках обязательного медицинского страхования.
Поликлиникой оказываются платные медицинские услуги как взрослым, так и детям независимо от
места жительства — на основании договоров на оказание платных медицинских услуг. Такой
договор для получения разовых медицинских услуг или на условиях длительного медицинского
обслуживания, в том числе по специальным программам (например, по программе ведения
беременности) можно заключить непосредственно в поликлинике при наличии документа,
удостоверяющего личность (паспорт). Кроме того, медицинские услуги можно получить по
программам добровольного медицинского страхования (ДМС), прикрепившись к поликлинике через
страховые медицинские организации. В зависимости от условий медицинского обслуживания при
наличии показаний оформляются листки нетрудоспособности.
Такой
договор для получения разовых медицинских услуг или на условиях длительного медицинского
обслуживания, в том числе по специальным программам (например, по программе ведения
беременности) можно заключить непосредственно в поликлинике при наличии документа,
удостоверяющего личность (паспорт). Кроме того, медицинские услуги можно получить по
программам добровольного медицинского страхования (ДМС), прикрепившись к поликлинике через
страховые медицинские организации. В зависимости от условий медицинского обслуживания при
наличии показаний оформляются листки нетрудоспособности.
(PDF) Эндокринные репродуктивные эффекты у моллюсков
Гиббс, PE, 2005. Дефект мужских половых органов (синдром Дамптона) у щенков
Nucella lapillus (Neogastropoda): предполагаемая менделевская наследственность, основанная на
экспериментах по разведению в лабораторных условиях. . J. Mar. Biol. Доц. СОЕДИНЕННОЕ КОРОЛЕВСТВО. 85, 143–150.
Гиббс П.Э., Паско П.Л., Берт Г.Р., 1988. Изменение пола у самки щенка,
Nucella lapillus, вызванное действием трибутилолова из точек против обрастания. J. Mar.Биол.
доц. СОЕДИНЕННОЕ КОРОЛЕВСТВО. 68, 715–731.
Гудинг, М.П., Уилсон, В.С., Фольмар, Л.С., Маркович, Д.Т., ЛеБлан, Г.А.,
2003. Биоцид трибутилолово снижает накопление тестостерона в виде
сложных эфиров жирных кислотв грязевой улитке (Ileta) obsoso. Environ. Здоровье
Персп. 111, 426–430.
Готфрид, Х., Дорфман, Р.И., 1970. Стероиды беспозвоночных V. Биосинтез стероидов in vitro
с помощью овотестиса мужской фазы слизняков (Ariolimax
californicus).Gen. Comp. Эндокринол. 15, 120–138.
Guillette Jr., LJ, Cross, TS, Masson, GR, Matter, JM, Percival, HF,
Woodward, AR, 1994. Аномалии развития гонад и
аномальных концентраций половых гормонов у молодых аллигаторов от заражения.
естественные и контролируемые озера во Флориде. Environ. Перспектива здоровья. 102, 680–688.
Guillette Jr., L.J., Pickford, D.B., Grain, D.A., Rooney, A.A., Percival, H.F.,
1996. Уменьшение размера полового члена и концентрации тестостерона в плазме у
молодых аллигаторов, живущих в загрязненной среде.Gen. Comp.
Эндокринол. 101, 32–42.
Хеллоу, Дж., Йейтс, П.С.С., Ганье, Ф., 2003. Химические загрязнители и биологические
индикаторов здоровья мидий во время гаметогенеза. Environ. Toxicol. Chem.
22, 2080–2087.
Henry, J., Favrel, P., Boucaud-Camou, E., 1997. Выделение и идентификация
нового пептида, родственного Ala-Pro-Gly-Trp-амиду, ингибирующего подвижность
зрелого яйцевода. у каракатиц Sepia officinalis.Пептиды 18,
1469–1474.
Хайнс, Г.А., Брайан, П.Дж., Уоссон, К.М., Макклинток, Дж. Б., Уоттс, С.А., 1996.
Метаболизм половых стероидов у антарктического птеропод Clione Antarctica
(Molluscka: Gastropoda). Инвертировать. Биол. 115, 113–119.
Хоригучи, Т., Тахигучи, Н., Чо, Х.С., Кодзима, М., Кая, М., Сираиси, Х.,
Морита, М., Хиросе, Х., Симидзу, М., 2000. Ово-семенники и нарушенный репродуктивный цикл
гигантского морского ушка Haliotis madaka: возможная связь
с загрязнением оловоорганическим веществом в месте сокращения популяции.Mar. Environ.
Рез. 50, 2236–229.
Хьюстон, Р.С., 1971. Репродуктивная биология Thais emarginata (Deshayes,
1839) и Thais canaliculata (Duclos, 1832). Велигер 13, 348–357.
Huet, M., Paulet, YM, Le Pennec, M., 1996. Выживание Nucella lapillus в загрязненной трибутилоловом районе
на западе Бретани: еще один пример дефекта мужских половых органов
(синдром Дамптона) в пользу выживание. Mar. Biol. 125, 543–549.
Джанер, Г., Lavado, R., Thibaut, R., Porte, C., 2005. Эффекты воздействия 17β-эстрадиола
на мидию Mytilus galloprovincialis: возможная регулирующая роль
для стероидных ацилтрансфераз. Акват. Toxicol. 75, 32–42.
Janer, G., Lyssimachou, A., Bachmann, J., Oehlmann, J., Schulte-Oehlmann, U.,
Porte, C., 2006. Половой диморфизм уровней этерифицированных стероидов у брюхоногих моллюсков
Marisa cornuarietis : действие ксеноандрогенных соединений. Стероиды 71,
435–444.
Джанер, Г., Порт, С., 2007. Половые стероиды и потенциальные механизмы негеномных
эндокринных нарушений у беспозвоночных. Экотоксикология 16, 145–160.
Джоблинг, С., Кейси, Д., Роджерс-Грей, Т., Оельманн, Дж., Шульте-Оельманн, У.,
Павловски, С., Баунбек, Т., Тернер, А.П., Тайлер, CR , 2003. Сравнительный ответ моллюсков и рыб
на эстрогены окружающей среды и эстрогенные сточные воды
. Акват. Toxicol. 65, 205–220.
Джоблинг, С., Нолан, М., Тайлер, К.Р., Брайти, Г., Самптер, Дж. П., 1998. Распространенное нарушение половой жизни
у диких рыб. Environ. Sci. Technol. 32, 2498–2506.
Кей, Дж., Бриджем, Дж. Т., Торнтон, Дж. У., 2006. Рецептор эстрогена
Octopus vulgaris является конститутивным активатором транскрипции: эволюционное и
функциональное значение. Эндокринология 147, 3861–3869.
Кетата, И., Гермази, Ф., Ребай, Т., Хамза-Чаффай, А., 2007a. Изменение концентраций стероидов
во время репродуктивного цикла моллюска Ruditapes
decussatus: годичное исследование в районе залива Габес.Комп. Биохим.
Physiol. А 147, 424–431.
Кетата, И., Смауи-Дамак, В., Гермази, Ф., Ребай, Т., Хамза-Чаффай, А.,
2007b. Эндокринные нарушения in situ кадмия в репродукции
Ruditapes decussatus. Комп. Biocem. Physiol. С 146, 415–430.
Кишида, М., Накао, Р., Новилло, А., Каллард, И.П., Осада, М., 2005. Молекулярное клонирование
и анализ экспрессии фрагментов кДНК, связанных с рецептором эстрогена
из синей мидии, Mytilus edulis .Proc.Jap.Soc.Comp.
Эндокринол. 20, 75.
Kluytmans, J.H., Brands, F., Zandee, D.I., 1988. Взаимодействие кадмия с репродуктивным циклом Mytilus edulis
. Mar. Environ. Res. 24, 189–192.
Labadie, P., Peck, M., Minier, C., Hill, E.M., 2007. Идентификация стероидных конъюгатов
сложных эфиров жирных кислот, образовавшихся in vivo у Mytilus edulis в результате воздействия
эстрогенов. Стероиды 72, 41–49.
Лафонт Р., 2000. Эндокринология беспозвоночных.Экотоксикология 9, 41–57.
Лафонт Р., Матье М., 2007. Стероиды у водных беспозвоночных. Экотоксикология
16, 109–130.
Lavado, R., Janer, J., Porte, C., 2006. Уровни стероидов и метаболизм стероидов в мидии Mytilus edulis
: модулирующий эффект диспергированной сырой нефти и
алкилфенолов. Акват. Toxicol. 78, S65 – S72.
ЛеБлан, Г.А., Кэмпбелл, П.М., ден Бестен, П., Браун, Р.П., Чанг, Е.С.,
Coats, J.R., de Fur, P.Л., Дхадиалла, Т., Эдвардс, Дж., Риддифорд, Л.М.,
Симпсон, М.Г., Снелл, Т.В., Торндайк, М., Мацумура, Ф., 1999. Эндокринология беспозвоночных
. В: deFur, P.L., Crane, M., Ingersoll, C.,
Tattersfield, L. (Eds.), Эндокринные нарушения у беспозвоночных: Endocrinol-
ogy, Testing and Assessment. SETAC Press, Пенсакола, Флорида, стр. 23–106.
Le Curieux-Belfond, O., Fievet, B., Séralini, G.E., Mathieu, M., 2005. Краткосрочная биоаккумуляция, циркуляция и метаболизм эстрадиола-17β в
oyster Crassostrea gigas.J. Exp. Mar. Biol. Ecol. 325, 125–133.
Le Curieux-Belfond, O., Moslemi, S., Mathieu, M., Séralini, GE, 2001.
Метаболизм андрогенов в устрице Crassostrea gigas: доказательства активности 17β-HSD
и характеристика активности, подобной ароматазе ингибируется
фармакологическими соединениями и загрязнителем морской среды. J. Steroid Biochem.
Мол. Биол. 78, 359–366.
Ли, Л.Дж., Гарден, Р.У., Флойд, П.Д., Мороз, Т.П., Глисон, Дж. М., Свидлер, Дж.V.,
Pasa-Tolic, L., Smith, R.D., 1999. Пептиды гормона яйцекладки в семействе
Aplysiidae. J. Exp. Биол. 202, 2961–2973.
Ли, К., Осада, М., Судзуки, Т., Мори, К., 1998. Изменения в вителлине во время оогенеза
и влияние эстрадиола на вителлогенез у тихоокеанских устриц
Crassostrea gigas. Инвертировать. Репрод. Dev. 33, 87–93.
Lubet, P., Mathieu, M., 1990. Les régulations endocriniennes chez les
mollusques bivalves.Аня. Биол. 29, 235–252.
Матье, М., 1994. Эндокринный контроль углеводного обмена у моллюсков.
Перспектива. Комп. Эндокринол. 471–474.
Матье, М., Роббинс, И., Любет, П., 1991. Нейроэндокринология Mytilus
edulis. Аквакультура. 94, 213–223.
Мацумото, Т., Накамура, А.М., Мори, К., Акияма, И., Хиросе, Х., Такахаши,
,Ю., 2007. Рецептор эстрогена устрицы: клонирование кДНК и иммунолокализация.
Gen. Comp.Эндокринол. 151, 195–201.
Мацумото, Т., Осада, М., Осава, Ю., Мори, К., 1997. Профиль гонадных эстрогенов
и иммуногистохимическая локализация стероидогенных ферментов в устрицах и гребешках
во время полового созревания. Комп. Биохим. Physiol. B
118, 811–817.
Matthiessen, P., Kilbride, R., Mason, C., Pendle, M., Rees, H., Waldock, R.,
1999. Мониторинг восстановления бентического сообщества в реке Крауч
после Загрязнение TBT.Отчет для Департамента окружающей среды, транспорта и регионов Великобритании
. Центр окружающей среды, рыболовства
и науки аквакультуры, Бернхэм-он-Крауч, Великобритания, стр. 51. EPG 1/5/92.
Маквей, М.Дж., Кук, Г.М., 2003. Ингибирование активности микросомальной 3β-
-гидроксистероиддегидрогеназы семенников крысы под действием трибутилолова. J. Steroid Biochem.
Мол. Биол. 86, 99–105.
Minier, C., Abarnou, A., Le Guellec, A.M., Jaouen-Madoulet, A., Tutundjian,
R., Bocquené, G., Leboulenger, F., 2006. Пилотное исследование мониторинга загрязнения
, включающее химический анализ и измерения биомаркеров в устье Сены
с использованием мидий зебры (Dreissena polymorpha). Environ. Toxicol.
Chem. 25, 112–119.
Minier, C., Caltot, G., Leboulenger, F., Hill, E.M., 2000. Расследование
случаев интерсекс-рыб в регионах Приморской Сены и Сассекса. Analusis
28, 801–806.
Морсилло, Ю., Alabalat, A., Porte, C., 1999. Мидии как стражи оловоорганического загрязнения
: биоаккумуляция и влияние на P450-опосредованную активность ароматазы.
Окружающая среда. Toxicol. Chem. 18, 1203–1208.
Morcillo, Y., Porte, C., 2000. Доказательства эндокринных нарушений у моллюсков
(Ruditapes decussata), пересаженных в загрязненную трибутилоловом среду.
Окружающая среда. Загрязнение. 107, 47–52.
268 I. Ketata et al. / Сравнительная биохимия и физиология, часть C 147 (2008) 261–270
Этот сайт использует файлы cookie для повышения производительности.Если ваш браузер не принимает файлы cookie, вы не можете просматривать этот сайт.
Настройка вашего браузера для приема файлов cookie
Существует множество причин, по которым cookie не может быть установлен правильно. Ниже приведены наиболее частые причины:
- В вашем браузере отключены файлы cookie. Вам необходимо сбросить настройки своего браузера, чтобы он принимал файлы cookie, или чтобы спросить вас, хотите ли вы принимать файлы cookie.
- Ваш браузер спрашивает вас, хотите ли вы принимать файлы cookie, и вы отказались.Чтобы принять файлы cookie с этого сайта, нажмите кнопку «Назад» и примите файлы cookie.
- Ваш браузер не поддерживает файлы cookie. Если вы подозреваете это, попробуйте другой браузер.
- Дата на вашем компьютере в прошлом. Если часы вашего компьютера показывают дату до 1 января 1970 г., браузер автоматически забудет файл cookie. Чтобы исправить это, установите правильное время и дату на своем компьютере.
- Вы установили приложение, которое отслеживает или блокирует установку файлов cookie.Вы должны отключить приложение при входе в систему или проконсультироваться с системным администратором.
Почему этому сайту требуются файлы cookie?
Этот сайт использует файлы cookie для повышения производительности, запоминая, что вы вошли в систему, когда переходите со страницы на страницу. Чтобы предоставить доступ без файлов cookie потребует, чтобы сайт создавал новый сеанс для каждой посещаемой страницы, что замедляет работу системы до неприемлемого уровня.
Что сохраняется в файле cookie?
Этот сайт не хранит ничего, кроме автоматически сгенерированного идентификатора сеанса в cookie; никакая другая информация не фиксируется.
Как правило, в файле cookie может храниться только информация, которую вы предоставляете, или выбор, который вы делаете при посещении веб-сайта. Например, сайт не может определить ваше имя электронной почты, пока вы не введете его. Разрешение веб-сайту создавать файлы cookie не дает этому или любому другому сайту доступа к остальной части вашего компьютера, и только сайт, который создал файл cookie, может его прочитать.
«Незаменимые минералы» | EURO VEDA Healing
«Незаменимые минералы» | ЕВРО ВЕДА ИсцелениеВы здесь:
- Home
- Общая информация
- Essential Minerals
«Основные минералы» с гарпагофитумом, бамбуком, Erigeron canadensis и раковиной моллюска
- Описание
- Harpagophytum (Коготь дьявола) — род растений из семейства кунжутных, произрастающих на юге Африки.Своим общим названием «Коготь дьявола» он обязан необычному внешнему виду плода с крючками. Крупные клубневые корни растения используются для уменьшения боли и лихорадки, а также для стимуляции пищеварения. Европейские колонисты принесли дьявольский коготь в Европу, где его использовали для лечения артрита.
- Бамбук — это цветущее многолетнее вечнозеленое растение семейства травянистых Poaceae. Бамбук — одно из самых быстрорастущих растений в мире. В Южной Азии, Юго-Восточной Азии и Восточной Азии бамбук используется в строительных материалах, как источник пищи и как универсальный сырьевой продукт.Качественный бамбук прочнее стали. В своем родном регионе бамбук использовался в медицине с древних времен.
- Erigeron Canadensis (Horseweed) — однолетнее растение, произрастающее на большей части территории Северной и Центральной Америки. Общие названия включают канадский водоросль. Его можно найти в полях, лугах и садах.
- Раковина моллюска представляет собой известковый экзоскелет, который охватывает, поддерживает и защищает мягкие части животного в типе моллюсков, который включает улиток, моллюсков, раковины клыков и несколько других классов.Не все моллюски с раковиной обитают в море, многие живут на суше и в пресной воде.
- Возможные достоинства
- Коготь дьявола обладает обезболивающими, седативными и мочегонными свойствами. Этот растительный препарат упоминается в Европейской фармакопее. Это часто используемый компонент ряда препаратов и пищевых добавок из-за его противоревматического действия. Дьявольский коготь используется при атеросклерозе, артрите, подагре, мышечной боли (миалгии), боли в спине, тендините, боли в груди, желудочно-кишечном расстройстве или изжоге, лихорадке и мигренозной головной боли.Он также используется при трудностях при родах, нарушениях менструального цикла, аллергических реакциях, потере аппетита и заболеваниях почек / мочевого пузыря
- Бамбук считается охлаждающим, успокаивающим и отводящим мокроту. Обстриженные молодые побеги, смола (жидкая и засохшая) и листья имеют лекарственную ценность, хотя их применение немного различается. Табашир (также называемый бамбуковой манной или бамбуковым кремнеземом) — одно из основных веществ из бамбука, используемых в аюрведической и тибетской медицине, поскольку он очень богат кремнеземом.Его свойства включают: стимулирующее, вяжущее, жаропонижающее, тонизирующее, спазмолитическое и афродизиакальное средство. Кремнезем помогает стимулировать синтез коллагена в костях, тканях и соединительной ткани и, таким образом, способствует реминерализации костей и суставов и способствует образованию хрящевой ткани. Это позволяет восстановить подвижность и уменьшить боли, связанные с заболеваниями суставов. Он помогает поддерживать эластичность стенок артерий и борется с атеросклерозом. Он также помогает поддерживать эластичность кожи и действует против морщин и растяжек.
- Конская водоросль — это традиционное североамериканское лекарственное средство на травах, которое раньше кипятили для приготовления пара для пота или принимали в качестве нюхательного табака, чтобы стимулировать чихание во время простуды. В настоящее время его больше всего ценят за терпкость, поскольку его используют при лечении желудочно-кишечных заболеваний, таких как диарея и дизентерия. Считается, что это очень эффективное средство от кровотечения при геморрое. Все растение обладает противоревматическим, вяжущим, бальзамическим, мочегонным, болеутоляющим, кровоостанавливающим, тонизирующим и глистогонным средством.Он особенно богат флавонами и дубильными веществами. Его традиционно используют для лечения ревматизма и воспалительных болей в суставах.
- Раковины моллюсков очень богаты кальцием и используются в фитотерапии для реминерализации.
- Я обычно прописываю состав «Незаменимые минералы» для укрепления костей и суставов; особенно
- Для лечения ревматических болей и болей в суставах , особенно остеоартрита и подагры
- Для лечения остеопороза и позвоночной грыжи диска
- Для поддержки наращивания костей и хрящей
- Рекомендуемое базовое лечение
- Принимать по 2 капсулы 3 раза в день, запивая большим стаканом воды перед едой (утром, в полдень, вечером)
- Essential Minerals не рекомендуется детям и лицам, получившим рану после травмы или операции.Каждое лечение должно проходить под наблюдением профессионального практикующего врача, особенно если пациент страдает желчными камнями или сердечно-сосудистыми заболеваниями.
- Диабетикам необходимо следить за уровнем сахара в крови, поскольку дьявольский коготь может снизить уровень глюкозы в крови.
- Действие кремнезема может усиливаться теми препаратами, которые разжижают кровь.
необычных режимов оогенеза и размножения двустворчатых моллюсков: случай Gaimardia trapesina (Mollusca: Gaimardiidae) в JSTOR
AbstractЯ описываю необычный случай фолликулярного оогенеза у двустворчатого моллюска Gaimardia trapesina, обычного морского двустворчатого моллюска из региона Магеллана и прилегающих субантарктических вод, члены которого несут потомство своих развивающихся личинок.Гонада у G. trapesina — это ацинусный орган, проникающий в перивисцеральную соединительную ткань; стенки ацинусов образованы высокими тонкими клетками с дистальными ядрами, поддерживаемыми тонким слоем соединительной ткани. В начале вителлогенеза каждый развивающийся ооцит окружается слоем фолликулярных клеток толщиной в одну клетку, которые могут происходить из стенки ацинуса. Клетки образуют фолликул, который полностью охватывает отдельные ооциты, за исключением базальной зоны, где ооциты контактируют со стенкой ацинуса.Фолликул сохраняется после окончания вителлогенеза и нереста. После высвобождения гамет персистирующий фолликул участвует в прикреплении яйцеклеток и развитии эмбрионов к межфиламентным соединениям внутренних и внешних полужаберных жабр, где зародыши инкубируются до тех пор, пока не вылупятся личинки педивелигеров на поздней стадии. Созревшие яйца имеют большие размеры (около 250 мкм в диаметре), что позволяет предположить, что развитие полностью лецитотрофное. Клетки фолликулов, которые опосредуют связи между развивающимися эмбрионами и особью матери, вероятно, имеют только механическую роль, обеспечивая поддержку и, возможно, облегчая аккомодацию большого количества эмбрионов, чтобы максимизировать жаберное пространство, доступное для вынашивания потомства.
Journal InformationInvertebrate Biology (IB) представляет оригинальные исследования и обзорные статьи по всем аспектам биологии беспозвоночных — морфологии и ультраструктуре; генетика, филогенетика и эволюция; физиология и экология; нейробиология, поведение и биомеханика; размножение и развитие; клеточная и молекулярная биология — и на всех типах беспозвоночных: простейших и многоклеточных, водных и наземных, свободноживущих и симбиотических. Все материалы проходят тщательную экспертную оценку.Журнал, издаваемый Американским микроскопическим обществом (AMS), входит в число старейших непрерывно публикуемых журналов в Соединенных Штатах и существует с 1880 года. В течение 100 лет (1895–1995) его название было «Труды Американского микроскопического общества»; с тех пор он был опубликован под названием «Биология беспозвоночных», чтобы отразить его нынешний широкий охват биологии беспозвоночных животных. Отдел зоологии беспозвоночных Общества интегративной и сравнительной биологии присоединяется к AMS в продвижении журнала, который публикуется совместно с Blackwell Publishing.
Информация для издателяWiley — глобальный поставщик решений для рабочих процессов с поддержкой контента в областях научных, технических, медицинских и научных исследований; профессиональное развитие; и образование. Наши основные направления деятельности выпускают научные, технические, медицинские и научные журналы, справочники, книги, услуги баз данных и рекламу; профессиональные книги, продукты по подписке, услуги по сертификации и обучению и онлайн-приложения; образовательный контент и услуги, включая интегрированные онлайн-ресурсы для преподавания и обучения для студентов и аспирантов, а также для учащихся на протяжении всей жизни.Основанная в 1807 году компания John Wiley & Sons, Inc. уже более 200 лет является ценным источником информации и понимания, помогая людям во всем мире удовлетворять их потребности и реализовывать их чаяния. Wiley опубликовал работы более 450 лауреатов Нобелевской премии во всех категориях: литература, экономика, физиология и медицина, физика, химия и мир. Wiley поддерживает партнерские отношения со многими ведущими мировыми обществами и ежегодно издает более 1500 рецензируемых журналов и более 1500 новых книг в печатном виде и в Интернете, а также базы данных, основные справочные материалы и лабораторные протоколы по предметам STMS.Благодаря растущему предложению открытого доступа, Wiley стремится к максимально широкому распространению и доступу к публикуемому контенту, а также поддерживает все устойчивые модели доступа. Наша онлайн-платформа, Wiley Online Library (wileyonlinelibrary.com), является одной из самых обширных в мире междисциплинарных коллекций онлайн-ресурсов, охватывающих жизнь, здоровье, социальные и физические науки и гуманитарные науки.
Жемчужная ферма на Пхукете, Таиланд
Жемчужная ферма на Пхукете
Хотите увидеть и узнать о процессе выращивания натурального жемчуга?
Жемчуг выращивают в 1 км от побережья залива Сапам, который расположен к востоку от Пхукета.Вы доберетесь туда на скоростном катере за 10 минут. Это индивидуальный тур. Здесь вы узнаете секреты рождения жемчуга. Климат восточного побережья Пхукета идеально подходит для выращивания жемчуга. Температура воды 26-29 ° С круглый год. Морские течения приносят и поддерживают запас питательных веществ для устриц.
Как выращивают жемчуг
Жемчуг Мабе. Когда устрица достигает 13 см в диаметре, бусинка имплантируется в мантию моллюска, и устрица растет еще 14 месяцев в естественной среде обитания.При имплантации бусинки очень легко повредить устрицу, поэтому на ферме работают только высококвалифицированные специалисты.
Жемчуг Акойя. Устрицы размером 5-10 см в диаметре собираются в море. Жемчужная раковина порождает жемчуг Акойя (Pinctada Fucada). Затем в соединительные ткани мантии моллюска имплантируется бусина. После имплантации моллюски проверяются в течение шести недель, и только по истечении этого времени их вывозят на ферму, где проводят еще 8-24 месяца.
Жемчуг Южных морей — самый крупный вид жемчуга. Для его выращивания используются очень редкие устрицы Pinctada Maxima. Бусины имплантируются в 24-месячные жемчужины и культивируются в течение 2-3 лет.
Во время экскурсии по Жемчужной ферме мы также предлагаем посетить уникальный магазин жемчуга — Аморн Жемчуг. Узнать больше о магазине и заказать трансфер можно здесь.
Немного истории
В 1967 году г-н Хун-Еанг Сэ-Тэу и г-жа Чун Тэ-Тэу приехали из Японии и открыли ферму по выращиванию натурального жемчуга.После многих лет напряженной работы семейный бизнес вырос до международного бренда — Phuket Pearl Industry Co Ltd.
Кремнистые отложения Южного озера Кантара, соленого озера в Южной Австралии.
Браун, Р.Г., 1965. Отложения в лагуне Коронг, Южная Австралия. Диссертация, Университет Аделаиды.
Деринг, Дж. А., 1986. Корреляция керна и общий приток наносов. В: B.E. Берглунд (ред.) Справочник по палеоэкологии и палеогидрологии голоцена .С. 247–272. Уайли, Чичестер.
Google Scholar
Деринг, Дж. А., 1991. Отложения озерных отложений отражают эрозионные процессы. Hydrobiologia 214 : 99–106.
Артикул Google Scholar
Де Деккер П. и Геддес М.С., 1980. Сезонная фауна соленых озер недалеко от лагуны Коронг, Южная Австралия. Австралийский журнал морских и пресноводных исследований 31 : 667–699.
Артикул Google Scholar
Эрлих, Р., 1983. Анализ размера не требует одежды, или моменты приходят и уходят? Журнал осадочной петрологии 53 : 1–12.
Google Scholar
Фридман Г.М., 1961. Различие между дюнным, пляжным и речным песками по текстурным характеристикам. Журнал осадочной петрологии 27 .514–529.
Google Scholar
Макманус, Дж., 1988. Определение и интерпретация размера зерен. В: М. Такер (ред.) Методы седиментологии . С. 63–85. Блэквелл, Оксфорд.
Google Scholar
Онгли, E.D., Брюн, M.C., и Персиваль, J.B., 1982. Физические и геохимические характеристики взвешенных твердых частиц, Уилтон-Крик, Онтарио. Hydrobiologia 91 : 41–57.
Google Scholar
Passega, R., 1964. Размер зерен, представленный C.M. узоры как геологический инструмент. Журнал осадочной петрологии 24 : 723–733.
Google Scholar
Петтиджон, Ф.Дж., 1975. Осадочные породы . Харпер и Роу, Нью-Йорк.
Google Scholar
Ройс, К.Ф., 1968. Распознавание речных сред по гранулометрическим характеристикам. Журнал осадочной петрологии 38 : 1171–1178.
Google Scholar
Слай П.Г., 1977 г. Осадочная среда в Великих озерах. В: H.L. Goltermann (Ed.) Proceedings of the SIL-UNESCO Symposium on Interactions between sashed and Freshwater . С. 76–82. Джанк, Гаага.
Google Scholar
Смит, Дж.Б. и Кершоу Р.С., 1979. Полевой справочник неморских моллюсков Юго-Восточной Австралии . Издательство Австралийского национального университета, Канберра.
Google Scholar
Стюарт, Х. Б., 1958. Отражения отложений в среде осадконакопления в лагуне Сан-Мигель, Нижняя Калифорния, Нью-Мексико. Бюллетень Американской ассоциации геологов-нефтяников 42 : 2567–2618.
CAS Google Scholar
Томас Р.Л., Кемп А.Л. и Льюис К.Ф.М., 1972. Поверхностные отложения озера Гурон. Канадский журнал наук о Земле 10 : 244–256.
Google Scholar
Томпсон Р., 1986. Палеомагнитное датирование. В: B.E. Берглунд (ред.) Справочник по палеоэкологии и палеогидрологии голоцена .С. 313–328. Уайли, Чичестер.
Google Scholar
Ван Уэвер, А., Стоун, А.В., 1980. Выделение субсреды отложения отложений в резервуарах с использованием параметров гранулометрического состава. Лимнологическое общество Южной Африки 6 (1): 59–65.
Google Scholar
фон дер Борт, Калифорния, 1976.Стратиграфия и формирование голоценовых доломитовых карбонатных отложений района Коронг, Южная Австралия. Журнал осадочной петрологии 46 : 952–966.
Google Scholar
Уоррен, Дж. К., 1988. Седиментология доломита Коронг в районе Солт-Крик, Южная Австралия. Карбонаты и эвапориты 3 (2): 175–199.
Артикул Google Scholar
Уоррен, Дж.К., 1990. Седиментология и минералогия доломитовых озер Коронг, Южная Австралия. Журнал осадочной петрологии 60 (6): 843–858.
CAS Google Scholar
Уильямс П.Ф., Руст Б.Р., 1969. Седиментология переплетенной реки. Журнал осадочной петрологии 39 : 649–679.
Google Scholar
Williamsm, W.D., 1986. Daphniopsis pusilla , кладоцер австралийских соленых озер: признание и экологические данные для палеолимонологов. Палеогеография, палеоклиматология, палеоэкология 54 : 305–316.
Артикул Google Scholar
Уильямс В.Д., 1991. Микрокосмы (микроэкосистемы) соленых озер как метод исследования свойств экосистемы. Verhandlungen Internationale Vereinigung Limnologie 24 : 1134–1138.
Google Scholar
Уильямс У.Д. и Коккинн М.Дж., 1988. Водно-болотные угодья и водная засоленная среда . Исследовательский проект AWRC 84/160, Отчет о завершении. Департамент водных ресурсов и энергетики, Канберра.
Google Scholar
Уильямс В.Д. и Меллор М.В., 1991. Экология Coxiella (Mollusc, Gastropoda, Prosobranchia), улитки, эндемичной для соленых озер Австралии. Палеогеография, палеоклиматология, палеоэкология 84 : 339–355.
Артикул Google Scholar
Эндокринные репродуктивные эффекты у моллюсков
Доступно на сайте www.sciencedirect.comСравнительная биохимия и физиология, часть C 147 (2008) 261–270 www.elsevier.com/locate/cbpc
Review
Репродуктивные эффекты, связанные с эндокринной системой моллюсков Имен Кетата а, Ксавье Денье b, Amel Hamza-Chaffai a, Christophe Minier b, ⁎ a
b
Ecotoxicologie Marine, UR 09-03, Université de Sfax, IPEIS, BP 805, 3018 Sfax, Тунисская лаборатория экотоксикологии, UPRES-EA 3222, Université du Havre, BP 540, 76058, Le Havre, France Получено 30 июля 2007 г .; получено в доработке 4 ноября 2007 г .; принято 26 ноября 2007 г. Доступно онлайн 4 декабря 2007 г.
Резюме Исследования эндокринных нарушений были главной темой последнего десятилетия.Хотя большинство исследований сосредоточено на позвоночных, беспозвоночные сейчас привлекают все больше внимания. В частности, данных о моллюсках становится все больше. Одним из наиболее задокументированных и наиболее актуальных примеров эндокринных нарушений является феномен импосекса, поражающий некоторые виды брюхоногих моллюсков. Но растущий интерес также связан с тем, что моллюски, особенно двустворчатые, являются хорошими биоиндикаторами, которые десятилетиями использовались в исследованиях окружающей среды, и что был достигнут прогресс в понимании физиологии и эндокринологии некоторых видов моллюсков.Недавние результаты предполагают, что на моллюсков могут отрицательно влиять соединения, которые изменяют их воспроизводство, и что метаболизм половых стероидов позвоночного типа или механизм действия могут быть вовлечены в эти эффекты. Тем не менее, эндокринная система моллюсков, по-видимому, во многих аспектах отличается от таковой у позвоночных, и половые стероиды могут не иметь одинакового значения для всех видов моллюсков. Это разнообразие представляет собой важную возможность изучить и понять новые и альтернативные механизмы эндокринных нарушений.© 2007 Elsevier Inc. Все права защищены. Ключевые слова: эндокринные разрушители; Эндокринная система; Гормоны; Моллюски; Репродукция
Содержание 1. 2. 3.
Введение. . . . . . . . . . . . . . . . . . . Эндокринная система моллюсков. . . . . . . . Влияние на размножение моллюсков. . . . . . . 3.1. Импосекс и интерсекс. . . . . . . . . . . 3.2. Производство яиц и эмбрионов. . . . . . . 3.3. Прочие эффекты. . . . . . . . . . . . . . . . 4. Механизмы действия эндокринных нарушений.. 4.1. Механизмы импосекса. . . . . . . . . . 4.2. Другие механизмы эндокринных нарушений 5. Выводы. . . . . . . . . . . . . . . . . . . Благодарности. . . . . . . . . . . . . . . . . . Рекомендации . . . . . . . . . . . . . . . . . . . . . . .
. . . . . . . . . . . .
. . . . . . . . . . . .
. . . . . . . . . . . .
. . . . . . . . . . . .
. . . . . . . . . . . .
. . . . . . . . . . . .
. . . .. . . . . . . .
. . . . . . . . . . . .
. . . . . . . . . . . .
. . . . . . . . . . . .
. . . . . . . . . . . .
. . . . . . . . . . . .
. . . . . . . . . . . .
. . . . . . . . . . . .
. . . . . . . . . . . .
. . . . . . . . . . . .
. . . . . . . . . . . .
. . . . . . . . . . . .
. . . . . . . . . . . .
. . . . . . . . . . . .
.. . . . . . . . . . .
. . . . . . . . . . . .
⁎ Автор для переписки. Лаборатория экотоксикологии, Гаврский университет, BP540, 76058 Гавр, Франция. Адрес электронной почты: [электронная почта защищена] (К. Минье). 1532-0456 / $ — см. Титульный лист © 2007 Elsevier Inc. Все права защищены. DOI: 10.1016 / j.cbpc.2007.11.007
. . . . . . . . . . . .
. . . . . . . . . . . .
. . . . . . . . . . . .
. . . . . . . . . . . .
. . . . . .. . . . . .
. . . . . . . . . . . .
. . . . . . . . . . . .
. . . . . . . . . . . .
. . . . . . . . . . . .
. . . . . . . . . . . .
. . . . . . . . . . . .
. . . . . . . . . . . .
. . . . . . . . . . . .
. . . . . . . . . . . .
. . . . . . . . . . . .
. . . . . . . . . . . .
. . . . . . . . . . . .
. . . . . . . . . . . .
.. . . . . . . . . . .
. . . . . . . . . . . .
262262 263263264 264 265 265 265 266 266 267
262
I. Ketata et al. / Сравнительная биохимия и физиология, часть C 147 (2008) 261–270
1. Введение В последние годы эндокринные нарушения в дикой природе были предметом многих обзоров и семинаров. Полевые наблюдения за аномальными репродуктивными организмами и сокращением популяций на загрязненных участках стимулировали серьезные исследовательские усилия, направленные на понимание связи между загрязнением окружающей среды и проблемами со здоровьем.Лабораторные исследования показали, что соединения могут влиять на синтез, секрецию, транспорт, метаболизм, механизм действия или клиренс естественных гормонов, ответственных за поддержание гомеостаза и регуляцию процессов развития (EPA, 1997). Эти химические вещества, таким образом, определяются как соединения, разрушающие эндокринную систему (EDC), и Европейское сообщество составило список из более чем 500 известных или предполагаемых EDC (EEC, 2001). Обеспокоенность здоровьем человека побудила к интенсивным исследованиям EDC.Сообщалось о снижении качества спермы, увеличении частоты аномалий развития мужских половых путей, преждевременном половом созревании, изменении развития нейронов, увеличении частоты опухолей молочных желез, простаты и яичек (обзор см. В ВОЗ, 2002). Это привело к многочисленным новым исследованиям видов позвоночных и к пересмотру уже сообщенных воздействий загрязняющих веществ на дикую природу. В сильно загрязненных местах репродуктивная недостаточность птиц, подвергшихся воздействию сельскохозяйственных и промышленных отходов (Fry and Toone, 1981), аномалии репродуктивных органов аллигаторов и белых медведей (Guillette et al., 1994, 1996; Wiig et al., 1998) и изменения пола ревертирующих рыб, подвергшихся воздействию эстрогенных стоков (Jobling et al., 1998, Minier et al., 2000). До недавнего времени экотоксикологи совершенно игнорировали воздействие загрязняющих веществ на представителей типа беспозвоночных. Ограниченное понимание нормальных эндокринных процессов у беспозвоночных затрудняло оценку химических эндокринных нарушений в полевых условиях (LeBlanc et al., 1999). Тем не менее беспозвоночные заслуживают большего внимания.Они являются ключевыми структурными и функциональными компонентами экосистемы, поскольку составляют 95% всех видов наземных и водных животных (Barnes, 1987; Wilson, 1999). Они не только являются источником пищи для многих позвоночных и трофическим звеном во многих экосистемах, они также играют важную роль в биогеохимическом круговороте (например, переработка детрита как в водной, так и в наземной среде) и в качестве инженеров экосистем. Это разнообразие проявляется в их эндокринной системе, разнообразии анатомических и физиологических особенностей, которые могут привести к уникальной восприимчивости к EDC.В отличие от позвоночных, беспозвоночные проявляют как бесполое, так и половое размножение, при этом 99% видов в какой-то момент своего жизненного цикла размножаются половым путем (Barnes et al., 1993). Кроме того, гермафродитизм и последовательный гермафродитизм (протандрия и протогиния) также широко распространены в основных таксонах беспозвоночных как нормальные формы сексуальности. Это разнообразие представляет собой важную возможность изучить и понять возможные механизмы эндокринных нарушений. Среди беспозвоночных моллюски являются предметом особого внимания исследований, что отражает их изобилие в природе, коммерческое и экологическое значение и их использование в качестве биоиндикаторов при мониторинге загрязнения воды (Philips, 1978; Minier et al., 2006).
Oetken et al. (2004) сообщили, что исследования видов моллюсков составили 55% исследований, проведенных по изучению эффектов EDC у беспозвоночных в 2003 году, тогда как в 1998 году они составили лишь 3,6%. Были предприняты важные усилия для лучшего понимания эндокринологии этого типа, хотя наши знания все еще фрагментарно. Однако это необходимый шаг для того, чтобы продемонстрировать какой-либо эффект нарушения эндокринной системы. В соответствии с этим фактом в данной статье делается попытка кратко и критически обобщить современные знания по репродуктивной эндокринологии моллюсков (более подробный обзор эндокринной системы см. В Lafont, 2000; deFur et al., 1999), а затем изучить основные сообщаемые эффекты нарушения эндокринной системы и связанные с ними механизмы. 2. Эндокринная система моллюсков. Эндокринная система включает химические посредники, которые секретируются для координации различных механизмов и физиологических процессов многоклеточного организма. Эта система позволяет организму адекватно реагировать на многочисленные внешние (окружающие) и внутренние (физиологические) сигналы. Нервная система играет центральную роль в этом координирующем процессе и усложняется в ходе эволюции.Нейросекреторные клетки были идентифицированы у диплобластных видов, где они присутствуют в сети нервных клеток. У моллюсков более организована нервная система и наблюдаются ганглии головного мозга и вентральная нервная цепь. В этой структуре нейросекреторные клетки присутствуют либо в нейрогемных органах, которые содержат нейросекреторные клетки и нервные клетки, либо в настоящих эндокринных железах (церебральных, плевральных, педальных и брюшных). Принято считать, что эндокринные железы первыми появились у видов моллюсков, хотя эндокринные клетки обнаружены у кольчатых червей, и что определение эндокринных желез может различаться у разных биологов.Эндокринную систему обычно описывают как систему, включающую несколько органов и химических медиаторов, действующих каскадом и, таким образом, подвергающихся множеству регуляторных процессов. Соответственно, системы контроля первого, второго и третьего порядка определяются в соответствии с количеством эндокринных желез и тканей-мишеней (Lafont, 2000). У моллюсков эндокринная система и особенно репродуктивный топор обычно включают только нейросекреторные клетки и другие эндокринные железы, такие как гонады, а регуляторная система третьего порядка была описана только для головоногих моллюсков (Di Cristo and Di Cosmo, 2007).В других группах многие из известных нейрогормонов действуют непосредственно на ткани-мишени, и только у видов брюхоногих существует достаточно доказательств того, что другая эндокринная железа, такая как гонада, выделяет химический посредник, который может действовать на другие ткани-мишени. У низших моллюсков еще предстоит продемонстрировать даже второй порядок регуляции. Эндокринную систему моллюсков можно рассматривать как наиболее разнообразную гормональную систему среди типов беспозвоночных. Он различается среди различных классов моллюсков и даже среди основной группы брюхоногих моллюсков, что отражает значительные различия в морфологии и истории жизни.Головоногие и большинство двустворчатых моллюсков являются гонохористами, тогда как большинство брюхоногих моллюсков являются одновременными или последовательными гермафродитами. Различия обнаружены в нейросекреторных органах, а также в гормонах
I. Ketata et al. / Сравнительная биохимия и физиология, часть C 147 (2008) 261–270
синтезируется организмами. Эти химические посланники можно разделить на две основные категории, учитывая их химические свойства и способ действия. Пептидные мессенджеры на сегодняшний день являются наиболее распространенными гормонами среди филума (и за его пределами среди типов беспозвоночных).Они секретируются нейросекреторными клетками или органами и действуют на мембранные рецепторы, а затем запускают каскад внутриклеточных событий, в конечном итоге приводящих к транскрипции генов. Примеры нейрогормонов моллюсков, участвующих в репродуктивном процессе, включают APGWamide, гормон каудодорсальных клеток (CDCH), гормоны спинного тела (DBH), гормон яйцекладки (ELH) и моллюскано-инсулиноподобные пептиды (MIPs) (Nagle et al., 1989; Гераертс и др., 1988; Смит и др., 1996; Ди Космо и Ди Кристо, 2006).Хотя нейроэндокринная система была особенно изучена у видов брюхоногих моллюсков, таких как Lymnaea, нет никаких сомнений в том, что пептиды играют роль в эндокринной системе других моллюсков, таких как двустворчатые моллюски. Например, рецепторы для GnRH или GnRH-подобных пептидов были обнаружены в гонаде двустворчатого моллюска Crassostrea gigas, которые, по-видимому, экспрессируются только во время развития гонад, поскольку в период сексуального покоя влияние этих нейрогормонов не регистрируется (Pazos and Mathieu, 1999). . Другие нейрогормоны производятся нервными ганглиями (церебральными, висцеральными или педальными).К ним относятся нейропептиды и аминергические нейросекреции (дофамин, норадреналин, серотонин). Стимулирующее действие нейрогормонов на половое созревание было показано на таких моллюсках, как Mytilus edulis (Mathieu et al., 1991) и C. gigas (Pazos and Mathieu, 1999). Нейрогормоны активируют как размножение гамет, так и вителлогенез, а также контролируют механизмы хранения энергии и процесс нереста (Lubet and Mathieu, 1990; Mathieu, 1994). Моллюски являются особенными среди протостомов, поскольку они считаются единственным филумом, который способен de novo синтезировать стероидные гормоны позвоночного типа, которые, в свою очередь, могут выполнять определенные физиологические роли.Тем не менее, следует отметить, что, хотя предполагается, что стероидные гормоны синтезируются из холестерина, фермент расщепления боковой цепи холестерина, то есть ключевой фермент, хорошо описанный у позвоночных, не был идентифицирован у моллюсков, и его активность описана только в слаге Ariolimax. californicus с очень низкой эффективностью (Gottfried, Dorfman, 1970). Эндогенное происхождение стероидов позвоночного типа у моллюсков вызывает сомнения, если принять во внимание их способность биоконцентрировать липофильные соединения в своих тканях (Le Curieux-Belfond et al., 2001, 2005; Labadie et al., 2007; Пек и др., 2007). Точно так же экдистероиды и терпеноиды были зарегистрированы в тканях моллюсков, хотя существует меньше доказательств их физиологической роли (ReisHenriques et al., 1990; Hines et al., 1996). Простагландины также могут поступать из окружающей воды и участвовать в воспроизводстве нескольких видов моллюсков, особенно за счет стимулирования нереста и пролиферации гамет (Freas, Grollman, 1981; Ono et al., 1982; Morse, 1984). Отсутствие эндогенного происхождения не означает, что данная молекула имеет биологическую роль (о чем свидетельствует витамин D у человека), и очевидно, что моллюски обладают активными ферментами, которые могут метаболизировать стероиды (De Longcamp et al., 1974; Гиббс и др., 1988; Ронис и Мейсон, 1996; Карро и Дросдровски,
263
1977). Во многих сообщениях подчеркивается влияние стероидов на половую дифференциацию, рост гонад, гаметогенез, яйценоскость, фертильность моллюсков, таким образом предполагая различные роли в воспроизводстве (Mori, 1969; Reis-Henriques and Coimbra, 1990; Osada et al., 1992; D ‘ Aniello et al., 1996; Matsumoto et al., 1997; Pazos and Mathieu, 1999; Wang and Croll, 2006). Примечательно, что некоторые эффекты могут показаться противоречивыми.Сообщалось, что инъекция эстрадиола вызывает изменение половой жизни от самцов к самкам у C. gigas (Mori, 1969), в то время как она увеличивает соотношение самцов и самок у Placopecten magellanicus (Wang and Croll, 2004). Более того, часто не сообщается о различиях в концентрациях половых стероидов между самцами и самками моллюсков, хотя в некоторых случаях годовые колебания могут казаться зависящими от пола (ReisHenriques et al., 1990; Gauthier-Clerc et al., 2006; Ketata et al., 2007a; Janer and Porte, 2007). Как наблюдается у позвоночных, стероидная активность может опосредоваться специфическими рецепторами.Ортологи эстрогеновых рецепторов обнаружены у представителей основных групп моллюсков: M. edulis (Kishida et al., 2005), C. gigas (Matsumoto et al., 2007), Aplysia californica (Thornton et al., 2003) и Octopus vulgaris (Keay et al., 2006). Однако исследования лигандов также показали, что рецепторы эстрогенов Octopus, Aplisia или Crassostrea не связывают эстрадиол или другие испытанные стероиды и, таким образом, не реагируют на эстрогены. Рецепторы являются конститутивными активаторами экспрессии генов из элементов ответа эстрогена (Thornton et al., 2003; Keay et al., 2006). Воздействие эстрадиола на мидии in vivo не привело к индукции рецептора эстрогена (Puinean et al., 2006). Таким образом, для эстрогенов могут существовать альтернативные механизмы действия (Stefano et al., 2003; Canesi et al., 2004), но еще предстоит выяснить. 3. Воздействие на размножение моллюсков Imposex считается лучшим и наиболее актуальным примером химически индуцированного эндокринного нарушения в дикой природе (ВОЗ, 2002). Тем не менее, помимо этого примера, уникального для переджаберных видов, количество других примеров эндокринных нарушений у моллюсков довольно ограничено.Можно утверждать, что это происходит из-за отсутствия у нас знаний об эндокринной системе моллюсков и, соответственно, недавно были получены новые данные, указывающие на то, что химическое воздействие на многие виды моллюсков может вызвать другие эффекты. 3.1. Импосекс и интерсекс Импосекс определяется как явление вирилизации или псевдогермафродитное состояние, характеризующееся развитием и наложением нефункциональных мужских половых признаков (половой член, семявыносящие протоки и / или семенные канальцы) у женщин.Nucella lapillus (Blaber, 1970), Ilyanassa obsolete (Smith, 1971), Thais emarginata (Houston, 1971) и Ocenebra erinacea (Féral, 1974) были первыми обнаруженными самками неогастропод, несущих пенис, но это явление широко распространено и затрагивает более 160 видов. из разных географических регионов (deFur et al., 1999; Shi et al., 2005). Imposex является специфической реакцией на оловоорганические соединения и вызывается почти исключительно трибутилоловом (TBT) в полевых условиях с эффективным водным порогом
264
I.Ketata et al. / Сравнительная биохимия и физиология, Часть C 147 (2008) 261–270
Концентрация близка к 1 нг TBT-Sn на литр. Интенсивность вирилизации у самок зависит от концентрации и в конечном итоге приводит к бесплодию. Эти эффекты имеют прямые последствия для репродуктивной способности популяции, ограничивая количество молодых особей и их вербовку во взрослую популяцию. Виды брюхоногих моллюсков, которые имеют низкую плодовитость, только ограниченную подвижность и вылупившиеся малорослые детеныши, страдают от сокращения или исчезновения популяции в сильно пораженных местах (Matthiessen et al., 1999). Однако высокое загрязнение TBT также может способствовать отбору афаллических особей N. lapillus (Гиббс, 1993, 2005; Huet et al., 1996). Эта генетически обусловленная резистентность, известная как синдром Дамптона (СД), снижает стерилизующий эффект импосекса, способствуя выживанию населения в районах, сильно загрязненных ТБТ. Не все виды переджаберных животных восприимчивы к импосексу, и межвидовые различия в восприимчивости изучены плохо. Самка барвинка Littorina littorea не развивает половой член в присутствии TBT, но демонстрирует при высоких концентрациях TBT (5–10 нг TBT-Sn / л) трансформацию его паллиальных органов в сторону мужской морфологической структуры (Bauer et al. ., 1995, 1997). Это явление называется интерсексом, и на более поздних стадиях у самок собачьих телохранителей развивается предстательная железа и бороздка для спермы, что приводит к ограничению их репродуктивных возможностей (Oehlmann et al., 1998). 3.2. Производство яиц и эмбрионов. Недавние исследования показали, что некоторые виды переджаберных животных могут быть очень чувствительны к химическим соединениям, известным как ксеноэстрогены, в исследованиях с использованием позвоночных. Oehlmann et al. (2000) сообщили о дозозависимом увеличении количества яиц, производимых Marisa cornuarietis, подвергшихся воздействию 1–25 мкг / л бисфенола A (BPA) и октилфенола (OP) с последующим уменьшением до 100 мкг / л.Это привело к накоплению яичных масс в женских паллиальных железах (белок и капсульные железы), которые были увеличены, и к разрыву яйцевода у всех самок в экспериментальных группах BPA и OP, независимо от концентрации ксеноэстрогенов. В конечном итоге смерть женщины (15% независимо от дозы) наблюдалась с незначительным сдвигом соотношения полов в пользу мужчин. Гистопатологический анализ гонад обоих полов во время эксперимента не дал никаких указаний на то, что на спермиогенез или оогенез влиял BPA или OP в применяемом диапазоне концентраций.Такие же эффекты наблюдались в поколении F1 при рассмотрении гипертрофии белка и капсульных желез. Интересно, что 100 мкг BPA / л затем привели к фенотипу «парадоксального импосекса» (учитывая, что BPA является эстрогенным соединением). Аналогичным образом, для грязевой улитки Potamopyrgus antipodarum воздействие BPA (1–300 мкг / кг сухих влажных отложений в Duft et al., 2003 или 1-25 мкг / л в Jobling et al., 2003) привело к увеличению общее производство эмбрионов после воздействия от 2 до 6 недель. Этот эффект был обусловлен значительным увеличением количества неочищенных эмбрионов, в то время как производство нормальных зародышей с оболочкой не могло быть затронуто.Через 8 недель связь «доза-реакция» была потеряна. Для OP такие же эффекты (большее количество эмбрионов) наблюдались у иловой улитки с четкой зависимостью от дозы только через 4 недели при воздействии через осадок
(1–300 мкг / кг сухого веса, Duft et al., 2003). ) или через 7 недель, когда в воде присутствовал алкилфенол (1–25 мкг / л, Jobling et al., 2003). У N. lapillus оценивалась повышенная продуктивность эмбрионов, но без зависимости от дозы (Oehlmann et al., 2000). Кроме того, это не привело к разрыву яйцевода или увеличению смертности, но производство мужской спермы упало до трети от контроля, и было измерено уменьшение размера полового члена и предстательной железы независимо от дозы.Эффекты нонилфенола (NP) и этинилэстрадиола (EE2) менее очевидны. Duft et al. (2003) сообщили об увеличении продуцирования эмбрионов грязевой улитки после 8-недельного воздействия отложений, загрязненных 10 и 100 мкг NP / кг сухого веса, но не заметного изменения при 30 мкг NP / кг сухого веса. Czech et al. (2001) сообщили о снижении яйценоскости после 12-недельного воздействия 100 мкг НП / л на L. stagnalis. У P. antipodarum отложения, загрязненные 30 мкг этинилэстрадиола (EE2) / кг, приводили к увеличению продуцирования эмбрионов (Duft et al., 2003). У тех же видов воздействие 25 и 100 нг EE2 / л значительно увеличивало производство новых эмбрионов (Jobling et al., 2003). Segner et al. (2003) сообщили об увеличении яйценоскости и задержке вылупления L. stagnalis, подвергшегося воздействию 500 нг EE2 / л. У M. cornuarietis 500 нг EE2 / л не увеличивали яйценоскость, но вызывали зависящий от времени феномен импосекса, эффект, который отменялся добавлением 1,25 мг ципротенона ацетата / л (Tillmann et al., 2001). 3.3. Другие эффекты Было зарегистрировано множество различных эффектов у видов моллюсков, подвергшихся воздействию химических загрязнителей, включая некоторые известные или предполагаемые нарушители эндокринной системы позвоночных, но их эндокринный механизм действия требует дальнейшего изучения.Среди них гистологические изменения могут быть более актуальными и более распространенными среди всех групп моллюсков. Задержка оогенеза или сперматогенеза, некроз канальцев, воспалительная реакция (инфильтрация фагоцитарных гемоцитов) и пролиферация соединительных тканей — наиболее часто описываемые гистологические признаки (Kluytmans et al., 1988; Li et al., 1998; Teh et al., 1999; Tillmann et al., 2001; Binelli et al., 2001; Aarab et al., 2004; Ortiz-Zarragoitia and Cajaraville, 2005). Ovotestis также отмечен у гонохорических моллюсков, включая брюхоногих моллюсков (Huet et al., 1996; Horiguchi et al., 2000) и двустворчатых моллюсков (Nice et al., 2003; Smaoui-Damak et al., 2006; Ketata et al., 2007a, b). Неравномерное соотношение полов по отношению к самцам наблюдалось у моллюсков (M. arenaria) и мидий (M. edulis), контактировавших с загрязненными реками (Gagné et al., 2003; Hellou et al., 2003). Индукция или изменение вителлогенеза, возможно, может быть хорошим индикатором эндокринного эффекта, и об этом сообщалось у нескольких видов двустворчатых моллюсков (Blaise et al., 1999; Aarab et al., 2004). Повышение концентрации вителлогенина в гонаде мидии Elliptiocompanata может объяснить нарушение репродуктивных возможностей (Blaise et al., 2003). У моллюсков Mya arenaria, подвергшихся воздействию городских сточных вод, повышенное фосфорилирование белков было измерено с помощью щелочно-лабильного фосфатного анализа (Gagné et al., 2002), а сточные воды индуцировали экспрессию рецептора эстрадиола у брюхоногих моллюсков N. lapillus (Castro et al., 2007) .
I. Ketata et al. / Сравнительная биохимия и физиология, часть C 147 (2008) 261–270
Canesi et al. (2007) сообщили, что лечение эстрадиолом индуцировало различные ферменты, которые играют роль в окислительном стрессе.4. Механизмы действия эндокринных нарушений. По определению, соединения, нарушающие работу эндокринной системы, могут изменять репродуктивную функцию, влияя на синтез, секрецию, транспорт, метаболизм, механизм действия или клиренс естественных гормонов (EPA, 1997), и, соответственно, многие из этих механизмов были исследованы в моллюски. 4.1. Механизмы импосекса. Féral (1977, 1980) обнаружил, что у брюхоногих моллюсков O. erinacea только самки, у которых развивался пенис, синтезировали нейрогормон, также присутствующий в педальных ганглиях самцов.Впоследствии в ходе лабораторных экспериментов Фераль и Ле Галл (1982) показали, что воздействие ТБО может вызвать аномальное высвобождение этого нейрогормона, вызывающее рост полового члена у молодых особей и самок в течение 24 часов после воздействия. Соответственно, предполагается, что TBT, который накапливается предпочтительно в нервах и ганглиях (Bryan et al., 1993), действует как нейротоксин, который стимулирует высвобождение нейрогормона, называемого морфогенным фактором полового члена (PMF), и индуцирует развитие вторичных половых признаков у мужчин. Пептидный гормон APGWamide был предложен в качестве предполагаемого PMF у Ilyanassa obsoleta, и было показано, что он вызывает импосекс при такой низкой концентрации, как 4 × 10–16 моль / животное (Oberdörster and McClellan-Green, 2000, 2002).APGWamide высоко консервативен среди моллюсков, включая двустворчатые моллюски, брюхоногие и головоногие моллюски, и может играть роль как в воспроизводстве, так и в развитии (Smith et al., 1997; Henry et al., 1997; Fan et al., 1997; Di Cristo et al. , 2005). TBT-индуцированный импосекс может подавляться совместным воздействием антиандрогена ципротерона ацетата, некоторых ингибиторов ароматазы, таких как SH 489 (1-метил-1,4-андростадиен-3,17дион) и флавона, эстрадиола и к индуктору цитохрома P450 3-метилхолантеру (Bettin et al., 1996; Обердёрстер и Макклеллан-Грин, 2002; Santos et al., 2005). Это предполагает, что стероиды могут участвовать в фенотипе импосекса. Соответственно, в относительно высоких дозах тестостерон может вызывать импосекс у нескольких видов, таких как Hinia reticulate, Nassarius reticulate, M. cornuarietis и N. lapillus (Spooner et al., 1991, Bettin et al., 1996, Schulte-Oehlmann et al. , 1995). Сообщалось о повышенных уровнях тестостерона после установления вторичных половых признаков (Spooner et al., 1991; Беттин и др., 1996; Santos et al., 2005) и могут участвовать в поддержании импосекса (Oberdörster and McClellan-Green, 2000). Сообщалось о противоречивых результатах по уровням свободного эстрадиола. В некоторых сообщениях показано сильное снижение эстрадиола (не связанное с ингибированием активности ароматазы), что изменило соотношение свободный тестостерон / свободный эстрадиол в пользу андрогенов (Morcillo et al., 1999; Morcillo and Porte, 2000). В других исследованиях не наблюдалось значительных изменений в титрах свободного эстрадиола (Spooner et al., 1991; Беттин и др., 1996; Schulte-Oehlmann et al., 1995). TBT может также влиять на несколько ферментов, участвующих в синтезе или выведении стероидов. Это включает ароматазу CYP19 у некоторых видов
265
(Bettin et al., 1996; Santos et al., 2005), но не всех (Morcillo et al., 1999). TBT может ингибировать превращение тестостерона в этерифицированные или сульфатированные формы, что приводит к снижению метаболического выведения этого гормона. Как следствие, повышение уровня свободного тестостерона может быть связано с индукцией импосекса (Ronis and Mason, 1996; Gooding et al., 2003; Santos et al., 2005). Недавние исследования также продемонстрировали, что TBT может связываться с ретиноидным X рецептором с более сильным сродством, чем природный лиганд (9-цис-ретиноевая кислота) у брюхоногих моллюсков Thais clavigera (Nishikawa et al., 2004). Было показано, что у людей RXR влияет на биосинтез стероидов посредством модуляции ароматазы P450 (Mu et al., 2001). Тем не менее, при сравнении видов следует соблюдать осторожность. Ферменты, такие как 5α-редуктазы и гидроксистероиддегидрогеназы, сильно ингибируются в субклеточных фракциях, выделенных из тканей позвоночных, но, вероятно, не являются специфическими мишенями для оловоорганических соединений у беспозвоночных (Doering et al., 2002; Маквей и Кук, 2003 г.). Кроме того, ферменты, такие как ароматаза и сульфотрансферазы, обладают низкой активностью в отношении моллюсков даже по сравнению с другими видами беспозвоночных (Janer et al., 2005). 4.2. Другие механизмы эндокринного нарушения. Возникновение драматических эндокринных эффектов, связанных с активностью, синтезом или элиминацией стероидов у позвоночных, и появление подобных стероидов в тканях моллюсков побудило исследовать аналогичные механизмы эндокринного нарушения у беспозвоночных.Было проведено множество лабораторных экспериментов с такими соединениями, как ксеноэстрогены позвоночных или другими соединениями, предположительно разрушающими эндокринную систему, но лишь немногие из них смогли продемонстрировать эндокринный механизм действия. Тем не менее, эксперименты, показывающие усиление производства яиц или эмбрионов у некоторых брюхоногих моллюсков после воздействия бисфенола А и октилфенола, ясно указывают на то, что эти соединения мешают эндокринно-регулируемому механизму. Чувствительность этих моллюсков к ксеноэстрогенам можно сравнить с позвоночными (Jobling et al., 2003). О влиянии модельного эстрогена 17β-эстрадиола на продукцию яиц или эмбрионов еще не сообщалось. Иногда было показано, что уровни стероидов в гонадах или в целом организме изменяются у нескольких видов, подвергшихся воздействию ксенобиотиков (Morcillo et al., 1998; Siah et al., 2003), но остается неясным, было ли это связано со специфическим действием на эндокринную систему. или результат непрямого токсического действия. Следует отметить, что концентрация стероидов в гонаде может сильно различаться у разных видов. Например, Janer et al.(2006) сообщили, что общая концентрация E2 составляла 5 и 25 нг / г сырого веса, соответственно, у самцов и самок брюхоногих моллюсков M. cornuarietis, тогда как у двустворчатых моллюсков M. arenaria концентрация E2 составляла от 0,15 до 0,4 нг / г сырого веса. для обоих полов (Gauthier-Clerc et al., 2006) и ниже 0,04 нг / г сырого веса M. edulis (Reis-Henriques et al., 1990; Puinean et al., 2006). В своем обзоре Janer and Porte (2007) сообщили, что уровни E2 в моллюсках были распределены на 4 порядка в зависимости от вида и метода, используемого для оценки уровней стероидов.Кроме того, моллюски могут в значительной степени концентрировать экзогенные стероиды, но, по-видимому, их количество составляет
266
I. Ketata et al. / Comparative Biochemistry and Physiology, Part C 147 (2008) 261–270
способна поддерживать концентрацию свободных стероидов в гонадах неизменной, даже если это влияет на общий уровень стероидов (Lavado et al., 2006; Puinean et al., 2006; Janer et al., 2006; Labadie et al., 2007). Индукция синтеза вителлогенина (vtg) была изучена и предложена в качестве биомаркера воздействия эстрогена на моллюсков (Blaise et al., 1999). Но четких зависимостей «доза-эффект» по-прежнему нет. Однократная инъекция E2 (1–1000 нмоль / животное) индуцировала вителлогенин-подобные белки, определяемые щелочно-лабильными фосфатами, в течение 72 часов у M. arenaria и E.companata (Blaise et al., 1999; Gagné et al., 2001a) , б, 2005). Повышенная концентрация щелочно-лабильных фосфатов была измерена как в гемолимфе, так и в гонадах двустворчатых моллюсков. Напротив, с помощью гель-анализа не было измерено никаких vtg-подобных белков в гемолимфе Anodonta cygnea после аналогичной обработки 250 нмоль E2 / животное (Riffeser and Hock, 2002) или в гонаде E.Complanata вводили 1–100 нмоль E2 / животное или подвергали воздействию в течение 7 дней до 1–10 000 нмоль E2 / л (Won and Novillo, 2005). Не наблюдалось значительного изменения концентрации vtg гонад после 20 дней ежедневной инъекции (180 нмоль / животное) в устрице C. gigas (Li et al., 1998) или через 60 дней в гребешке, повторно вводимой P. yessoensis (каждые 10 дней) с 180 нмоль E2 / животное (Osada et al., 2003). Существенные отличия от контроля были достигнуты только через 40 и 90 дней продолжительного лечения соответственно.Однако Osada et al. (2003) сообщили об индукции синтеза вителлогенина (обнаруженного с помощью ELISA) в культивируемых фрагментах яичников, обработанных в течение 5 дней экстрактом церебральных и педальных ганглиев, который может содержать фактор, способствующий вителлогенезу. 5. Выводы. В последнее время были предприняты значительные усилия для дальнейшего документирования эндокринологии моллюсков и изучения предполагаемых эндокринных разрушительных эффектов загрязнения окружающей среды у этих видов. Хотя эндокринная система моллюсков в значительной степени контролируется действием пептидных гормонов, многочисленные исследования изучали наличие и активность стероидных гормонов у этих беспозвоночных.Доказательства наличия и способности метаболизировать стероиды с помощью различных ферментов были получены почти для всех групп моллюсков (Lafont and Mathieu, 2007). Тем не менее, эндогенное происхождение и физиологическая роль все еще сомнительны и требуют дальнейшего изучения. Несколько исследований импосекса ясно показали, что тестостерон играет роль в индукции или поддержании дополнительных мужских половых органов у самок брюхоногих моллюсков, но эффекты могут в первую очередь быть результатом взаимодействия TBT с нейросекреторными клетками (Féral and Le Gall, 1982; Oberdörster and McClellanGreen, 2002).Повышенное производство яиц или эмбрионов у M. cornuarietis, P. antipodarum, N. lapillus и Limnea stagnalis при воздействии октилфенола и бисфенола А также предполагает, что эти соединения, известные как ксеноэстрогены позвоночных, могут нарушать эндокринную систему моллюсков. Тем не менее, основной механизм до сих пор неизвестен. У позвоночных оба соединения активны через рецептор эстрогена, но этот механизм не может быть задействован у моллюсков, поскольку не было продемонстрировано никаких доказательств роли эндогенных стероидов в реакции моллюсков на откладывание яиц.Однако нейрогормоны, такие как гормон каудодорсальных клеток
(CDCH), могут стимулировать яйцекладку у Lymnaea и Aplysia (Roubos and Vanheumen, 1994; Li et al., 1999). Таким образом, оба основных эндокринных разрушительных эффекта, о которых сообщалось у моллюсков, указывают на то, что исследовательские усилия по выяснению основных механизмов не должны ограничиваться стероидными гормонами и их механизмом действия, но что следует исследовать весь эндокринный механизм, включая его регуляторные процессы. Тем не менее, присутствие стероидов и описанные эффекты у моллюсков интригуют.Были получены некоторые явно противоречивые результаты. Например, индукция vtg была успешной или неудачной после воздействия эстрадиола (Gagné et al., 2001a; Won and Novillo, 2005). В других исследованиях наблюдалась задержка переменной продолжительности до регистрации повышенного vtg (Li et al., 1998; Osada et al., 2003). Это указывает на то, что продукция vtg может регулироваться по-разному у моллюсков, чем у позвоночных. Эффект может зависеть от метаболизма стероидов, продолжительности воздействия и времени воздействия.Кроме того, эстрадиол не может быть основным эффектором наблюдаемой реакции. Сообщалось, что инъекция эстрадиола вызывает изменение половой жизни от самцов к самкам у C. gigas (Mori, 1969), в то время как андрогенные, эстрогенные и прогестатические стероиды увеличивают соотношение самцов и самок у P. magellanicus (Wang and Croll, 2004). Более того, часто не сообщается о различиях в концентрациях половых стероидов между самцами и самками моллюсков. Эти результаты предполагают, что либо стероиды не играют той же роли у моллюсков, как у позвоночных (то, что мы называем половыми стероидами, в основном получено из наших знаний об эндокринологии позвоночных), либо эти стероиды быстро превращаются в другие или инактивируются.Интересно, что сообщалось о парадоксальном импосексе, вызванном ксеноэстрогенами (Tillmann et al., 2001), эффект, который можно сравнить с парадоксальной феминизацией, наблюдаемой у рыб, получавших большое количество андрогенов (Rinchard et al., 1999; Örn et al., др., 2003). Знание концентраций стероидов в месте действия и механизма, с помощью которого стероиды могут запускать клеточный ответ, значительно расширило бы наше понимание эндокринологии моллюсков. В целом следует иметь в виду, что моллюски представляют собой группу со значительным разнообразием, что может отражаться на их эндокринной системе.Анатомические и физиологические признаки в значительной степени варьировались в зависимости от типа. Некоторые явно противоречивые результаты могут отражать это разнообразие между видами. Нарушение передачи сигналов нейропептидного гормона после воздействия TBT у брюхоногих моллюсков подчеркивает уникальность их эндокринной системы и их потенциал нарушения. Это разнообразие представляет собой важную возможность изучить и понять новые и альтернативные механизмы эндокринных нарушений. Благодарности Эта работа была выполнена в рамках программы AUF (Agence Universitaire de la Francophonie) (PCSI637).Авторы выражают благодарность Sfax University (Тунис), ENIS и IPEIS за их помощь. X. Denier и C. Minier были поддержаны в этом исследовании европейской исследовательской программой Interreg III (Анализ рисков, связанных с эндокринными нарушениями — RAED, контракт № 156).
I. Ketata et al. / Сравнительная биохимия и физиология, часть C 147 (2008) 261–270
Ссылки Aarab, N., Minier, C., Lemaire, S., Echehardt, U., Hansen, PD, Andersen, OK, Narbonne, JF, 2004. Биохимические и гистологические реакции у мидий (Mytilus edilus), подвергшихся воздействию нефти Северного моря и смеси нефти Северного моря и алкилфенола.Mar. Environ. Res. 25, 437–441. Барнс, Р. Д., 1987. Зоология беспозвоночных, 5-е изд. Сондерс, Филадельфия. Барнс, Р.С.К., Каллоу, П., Олив, П.Дж.У., Голдинг, Д.В., 1993. Беспозвоночные. Новый синтез, второе издание. Blackwell Science, Оксфорд, Великобритания. Bauer, B., Fioroni, P., Die, I., Liebe, S., Oehlmann, J., Stroben, E., Watermann, B., 1995. Влияние TBT на женские половые органы (Littorina littorea): Возможный индикатор загрязнения трибутилоловом. Hydrobiologia 309, 15–27. Бауэр, Б., Фиорони, П., Schulte-Oehlmann, U., Oehlmann, J., Kalbfus, W., 1997. Использование (Littorina littorea) для мониторинга эффекта трибутилолова (TBT) — результаты немецкого исследования TBT 1994: 1995 и лабораторных экспериментов. Environ. Загрязнение. 96, 299–309. Bettin, C., Oehlmann, J., Stroben, E., 1996. Импосекс, вызванный TBT, у морских новогастропод опосредуется повышением уровня андрогенов. Хельгол. Meeresunters. 50, 299–317. Бинелли, А., Баккетта, Р., Вайлати, Г., Галасси, С., Провини, А., 2001. Загрязнение ДДТ в озере Маджоре (Н.Италия) и влияние на нерест мидий-зебр. Chemosphere 45, 409–415. Blaber, S.J.M., 1970. Возникновение пенисовидного выроста позади правого щупальца у истощенных самок (Nucella lapillus, L.). Proc. Малакол. Soc. Лондон. 39, 231–233. Блез, К., Ганье, Ф., Пеллерин, Дж., Хансен, П.Д., 1999. Определение вителлогенин-подобных свойств в гемолимфе (Mya arenaria) (Сагеней-фьорд, Канада): потенциальный биомаркер эндокринных нарушений. Environ. Toxicol. 14, 455–465. Блез К., Ганье Ф., Салазар М., Салазар, С., Троттье, С., Хансен, П.Д., 2003. Экспериментально вызванная феминизация пресноводных мидий после длительного воздействия городских сточных вод. Fresenius Environ. Бык. 12, 8. Брайан, Г.В., Брайт, Д.А., Хаммерстон, Л.Г., Берт, Г.Р., 1993. Поглощение, распределение в тканях и метаболизм меченного 14С трибутилолова (ТБТ) у детенышей собаки (Nucella lapillus). J. Mar. Biol. Доц. СОЕДИНЕННОЕ КОРОЛЕВСТВО. 73, 889–912. Canesi, L., Ciacci, C., Betti, M., Lorusso, L.C., Marchi, B., Burattini, S., Falcieri, E., Gallo, G., 2004. Быстрые эффекты 17-бета-эстрадиола на передачу сигналов клеток и функцию гемоцитов (Mytilus). Gen. Comp. Эндокринол. 136, 58–71. Canesi, L., Borghi, C., Fabbri, R., Ciacci, C., Lorusso, L.C., Gallo, G., Vergani, L., 2007. Влияние 17β-эстрадиола на пищеварительную железу мидий. Gen. Comp. Эндокринол. 153, 40–46. Карро, С., Дросдовски, М., 1977. Биосинтез стероидов in vivo гонадой каракатицы (Sepia officinalis). Gen. Comp. Эндокринол. 33, 554–565. Кастро, L.F.C., Мело, К., Гийо, Р., Мендес, И., Кейрос, С., Лима, Д., РейсХенрикес, М.А., Сантос, М.М., 2007b. Рецептор эстрогена у брюхоногих моллюсков Nucella lapillus: Модуляция после воздействия эстрогенных стоков? Акват. Toxicol. 84, 465–468. Чех П., Вебер К., Дитрих Д. Р., 2001. Влияние веществ, модулирующих эндокринную систему, на репродуктивную функцию у гермафродитной улитки (Lymnaea stagnalis, L). Акват. Toxicol. 53, 103–114. Д’Аниелло, А., Ди Космо, А., Ди Кристо, К., Ассизи, Л., Ботте, В., Ди Фиоре, М.М., 1996. Появление половых стероидных гормонов и их связывающих белков в (Octopus vulgaris, лам).Биохим. Биофиз. Res. Commun. 227, 782–788. деФур, П.Л., Крейн, М., Ингерсолл, К., Таттерсфилд, Л., 1999. Эндокринные нарушения у беспозвоночных: эндокринология, тестирование и оценка. Общество экологической токсикологии и химии Press, Пенсакола, Флорида, США, стр. 303. Де Лонгкамп Д., Любет П., Дросдовский М., 1974. Биосинтез стероидов in vitro гонадой мидии (Mytilus edulis). Gen. Comp. Эндокринол. 22, 116–127. Ди Космо, А., Ди Кристо, К., 2006. Пептиды моллюсков и размножение.В: Кастин А. (Ред.), Справочник по биологически активным пептидам. Academic Press, Нью-Йорк.
267
Ди Кристо, К., Ди Космо, А., 2007. Нейропептидергический контроль яйцевидной железы осьминога. Пептиды 28, 163–168. Ди Кристо, К., Ван Миннен, Дж., Ди Космо, А., 2005. Присутствие APGWamide в Octopus vulgaris: возможная роль в репродуктивном поведении. Пептиды 26, 53–62. Деринг, Д.Д., Стекельброк, С., Деринг, Т., Клингмюллер, Д., 2002. Влияние бутилолова на активность 5α-редуктазы человека 1 и 2 типа.Стероиды 67, 859–867. Duft, M., Schulte-Oehlmann, U., Weltje, L., Tillmann, M., Oehlmann, J., 2003. Стимулированное продуцирование эмбрионов как параметр эстрогенного воздействия через отложения в пресноводном грязевом улитке (Potamopyrgus antipodarum). Акват. Toxicol. 64, 437–449. EEC, 2001. Стратегия сообщества в отношении эндокринных нарушителей — ряда веществ, предположительно влияющих на гормональные системы человека и дикой природы. COM 262. Брюссель. 46pp. EPA, 1997. Специальный отчет об эндокринных нарушениях окружающей среды: оценка и анализ последствий.(Отчет EPA № EPA / 630 / R-96/012) Агентство по охране окружающей среды США, Вашингтон, округ Колумбия. Fan, X., Croll, RP, Wu, B., Fang, L., Shen, Q., Painter, SD, Nagle, GT, 1997. Молекулярное клонирование кДНК, кодирующей нейропептиды APGWamide и церебральный пептид 1: локализация APGWamide-подобная иммунореактивность в центральной нервной системе и мужских репродуктивных органах аплизии. J. Comp. Neurol. 387, 23–62. Фераль, К., 1974. Сравнительный этюд популяций д’Оценебры erinacea (L.) de Granville et d’Arcachon.Халиотис 4, 129–134. Féral, C., 1977. Анализируйте экспериментальную морфогенез и цикл пениса, ле женского пола, д’Оценебры erinacea, espèce gonochorique. Халиотис 6, 267–271. Féral, C., 1980. Apparition de femelles à tractus génital mâle externe chez Ocenebra erinacea (L.), gonochorique mollusque gonochorique de la station de Granville: исследование факторов, контролирующих аномалию. C. R. Hebd. Séances Acad. Sci. Париж, Сер. Д 290, 1003–1006. Фераль К., Ле Галль С., 1982.Экспериментальная индукция по загрязнению марихуаны (трибутилэтилен), активная нейроэндокринная система, контролирующая морфогенез пениса, ле женского пола, д’Оценебры эринацеи (моллюски, prosobranche gonochorique). C. R. Hebd. Séances Acad. Sci. Париж, Сер. III 295, 627–630. Freas, W., Grollman, S., 1981. Поглощение и связывание простагландинов морскими двустворчатыми моллюсками Modiolus demissus. J. Exp. Zool. 216, 225–234. Фрай Д.М., Тоун К.К., 1981. ДДТ-индуцированная феминизация эмбрионов чаек. Science 213, 922–924. Ганье, Ф., André, C., Blaise, C., 2005. Повышенная экспрессия гена вителлогенина в мидии Elliptiocompanata, подвергшейся воздействию эстрадиола-17β. Fresenius Environ. Бык. 14, 861–866. Gagné, F., Blaise, C., Pellerin, J., Gauthier-Clerc, S., 2002. Изменение биохимических свойств женских гонад и желтков у моллюска Mya arenaria на загрязненных участках во фьорде Сагеней. Mar. Environ. Res. 53, 295–310. Ганье, Ф., Блез, К., Пеллерин, Дж., Пеллетье, Э., Дувиль, М., Готье-Клерк, С., Виглино, Л., 2003. Смена пола у моллюсков с мягким панцирем (Mya arenaria) в приливной зоне реки Святого Лаврентия (Квебек, Канада). Комп. Биохим. Physiol. B 134, 189–198. Ганье, Ф., Блез, К., Салазар, М., Салазар, С., Хансен, П.Д., 2001a. Оценка эстрогенного воздействия городских сточных вод на пресноводных мидий (Elliptio Complanata). Комп. Биохим. Physiol. С 128, 213–225. Ганье, Ф., Блез, К., Лашанс, Б., Сунахара, Г.И., Сабик, Х., 2001b. Доказательства эстрогенного воздействия копростанола на пресноводных мидий (Elliptio communanata).Environ. Загрязнение. 115, 97–106. Готье-Клерк, С., Пеллерин, Дж., Амиард, Дж. К., 2006. Концентрации эстрадиола-17β и тестостерона у мужчин и женщин Mya arenaria (Mollusca bivalvia) во время репродуктивного цикла. Gen. Comp. Эндокринол. 145, 133–139. Гераертс В.П.М., Вройгденхил Э., Эбберинк Р.Х.М., 1988. Биоактивные пептиды моллюсков. В: Thorndyle, M.C., Goldsworthy, G.J. (Ред.), Нейрогормоны у беспозвоночных. Издательство Кембриджского университета, Кембридж, Великобритания, стр. 261–281. Гиббс П.Е., 1993. Дефект мужских половых органов у щенка Nucella lapillus (Neogastropoda), способствующий выживанию в зоне, загрязненной ТБО.J. Mar. Biol. Доц. СОЕДИНЕННОЕ КОРОЛЕВСТВО. 73, 667–678.
268
I. Ketata et al. / Comparative Biochemistry and Physiology, Part C 147 (2008) 261–270
Gibbs, PE, 2005. Дефект мужских половых органов (синдром Дамптона) у щенка Nucella lapillus (Neogastropoda): Менделирующая наследственность сделана на основании экспериментов по разведению . J. Mar. Biol. Доц. СОЕДИНЕННОЕ КОРОЛЕВСТВО. 85, 143–150. Гиббс П.Е., Паско П.Л., Берт Г.Р., 1988. Изменение пола у самки щенков Nucella lapillus, вызванное действием трибутилолова из точек против обрастания.J. Mar. Biol. Доц. СОЕДИНЕННОЕ КОРОЛЕВСТВО. 68, 715–731. Гудинг, М.П., Уилсон, В.С., Фолмар, Л.С., Маркович, Д.Т., ЛеБлан, Г.А., 2003. Биоцид трибутилолово снижает накопление тестостерона в виде сложных эфиров жирных кислот в грязевой улитке (Ilyanassa obsoleta). Environ. Health Persp. 111, 426–430. Готфрид, Х., Дорфман, Р.И., 1970. Стероиды беспозвоночных V. Биосинтез стероидов in vitro посредством овотестиса мужской фазы слизняков (Ariolimax californicus). Gen. Comp. Эндокринол. 15, 120–138. Гийетт младший, Л.Дж., Кросс, Т.С., Массон, Г.Р., Маттер, Дж. М., Персиваль, Х.Ф., Вудворд, А.Р., 1994. Аномалии развития гонад и аномальные концентрации половых гормонов у молодых аллигаторов из загрязненных и контрольных озер Флориды. Environ. Перспектива здоровья. 102, 680–688. Гиллетт-младший, Л.Дж., Пикфорд, Д.Б., Грейн, Д.А., Руни, А.А., Персиваль, Х.Ф., 1996. Уменьшение размера полового члена и концентрации тестостерона в плазме у молодых аллигаторов, живущих в загрязненной среде. Gen. Comp. Эндокринол.101, 32–42. Хеллоу, Дж., Йейтс, П.С.С., Ганье, Ф., 2003. Химические загрязнители и биологические индикаторы здоровья мидий во время гаметогенеза. Environ. Toxicol. Chem. 22, 2080–2087. Henry, J., Favrel, P., Boucaud-Camou, E., 1997. Выделение и идентификация нового пептида, родственного Ala-Pro-Gly-Trp-амиду, ингибирующего подвижность зрелого яйцевода каракатиц, Sepia officinalis . Пептиды 18, 1469–1474. Хайнс, Г.А., Брайан, П.Дж., Уоссон, К.М., МакКлинток, Дж. Б., Уоттс, С.А., 1996. Метаболизм половых стероидов у антарктического птеропод Clione Antarctica (Molluscka: Gastropoda).Инвертировать. Биол. 115, 113–119. Хоригучи, Т., Тахигучи, Н., Чо, Х.С., Кодзима, М., Кая, М., Сираиси, Х., Морита, М., Хиросе, Х., Симидзу, М., 2000. Ово-яички и нарушение репродуктивного цикла гигантского морского морского ушка Haliotis madaka: возможная связь с оловоорганическим загрязнением в месте сокращения популяции. Mar. Environ. Res. 50, 2236–229. Хьюстон, Р.С., 1971. Репродуктивная биология Thais emarginata (Deshayes, 1839) и Thais canaliculata (Duclos, 1832). Велигер 13, 348–357. Юэ М., Поле Ю.М., Ле Пеннек, М., 1996. Выживание Nucella lapillus в загрязненном трибутилоловом районе в западной Бретани: еще один пример мужского генитального дефекта (синдром Дамптона), способствующего выживанию. Mar. Biol. 125, 543–549. Джанер, Г., Лавадо, Р., Тибо, Р., Порте, С., 2005. Эффекты воздействия 17β-эстрадиола на мидии Mytilus galloprovincialis: возможная регулирующая роль стероидных ацилтрансфераз. Акват. Toxicol. 75, 32–42. Янер, Г., Лиссимаху, А., Бахманн, Дж., Оельманн, Дж., Шульте-Оельманн, У., Порт, К., 2006. Половой диморфизм уровней этерифицированных стероидов у брюхоногих моллюсков Marisa cornuarietis: влияние ксеноандрогенных соединений. Стероиды 71, 435–444. Джанер, Г., Порт, С., 2007. Половые стероиды и потенциальные механизмы негеномных эндокринных нарушений у беспозвоночных. Экотоксикология 16, 145–160. Джоблинг, С., Кейси, Д., Роджерс-Грей, Т., Оельманн, Дж., Шульте-Оельманн, У., Павловски, С., Баунбек, Т., Тернер, А.П., Тайлер, CR, 2003. Сравнительный реакция моллюсков и рыб на эстрогены окружающей среды и эстрогенные сточные воды.Акват. Toxicol. 65, 205–220. Джоблинг, С., Нолан, М., Тайлер, К.Р., Брайти, Г., Самптер, Дж. П., 1998. Широко распространенные сексуальные нарушения у диких рыб. Environ. Sci. Technol. 32, 2498–2506. Кей, Дж., Бриджем, Дж. Т., Торнтон, Дж. В., 2006. Рецептор эстрогена Octopus vulgaris является конститутивным активатором транскрипции: эволюционное и функциональное значение. Эндокринология 147, 3861–3869. Кетата, И., Гермази, Ф., Ребай, Т., Хамза-Чаффай, А., 2007a. Изменение концентраций стероидов во время репродуктивного цикла моллюска Ruditapes decussatus: однолетнее исследование в районе залива Габес.Комп. Биохим. Physiol. А 147, 424–431. Кетата, И., Смауи-Дамак, В., Гермази, Ф., Ребай, Т., Хамза-Чаффай, А., 2007b. Эффекты кадмия, нарушающие эндокринную систему in situ, на размножение Ruditapes decussatus. Комп. Biocem. Physiol. С 146, 415–430. Кишида, М., Накао, Р., Новилло, А., Каллард, И.П., Осада, М., 2005. Молекулярное клонирование и анализ экспрессии фрагментов кДНК, связанных с рецептором эстрогена
из голубой мидии, Mytilus edulis. Proc. Яп. Soc. Комп. Эндокринол. 20, 75. Kluytmans, J.H., Brands, F., Zandee, D.I., 1988. Взаимодействие кадмия с репродуктивным циклом Mytilus edulis. Mar. Environ. Res. 24, 189–192. Labadie, P., Peck, M., Minier, C., Hill, E.M., 2007. Идентификация конъюгатов стероидных эфиров жирных кислот, образующихся in vivo у Mytilus edulis в результате воздействия эстрогенов. Стероиды 72, 41–49. Лафон, Р., 2000. Эндокринология беспозвоночных. Экотоксикология 9, 41–57. Лафон, Р., Матье, М., 2007. Стероиды у водных беспозвоночных. Экотоксикология 16, 109–130.Lavado, R., Janer, J., Porte, C., 2006. Уровни стероидов и метаболизм стероидов в мидии Mytilus edulis: модулирующий эффект диспергированной сырой нефти и алкилфенолов. Акват. Toxicol. 78, S65 – S72. ЛеБлан, Джорджия, Кэмпбелл, П.М., ден Бестен, П., Браун, Р.П., Чанг, Э.С., Пальто, Дж. Р., де Фур, П.Л., Дхадиалла, Т., Эдвардс, Дж., Риддифорд, Л.М., Симпсон, М.Г., Снелл , Т.В., Торндайк, М., Мацумура, Ф., 1999. Эндокринология беспозвоночных. В: deFur, P.L., Crane, M., Ingersoll, C., Tattersfield, L.(Ред.), Эндокринные нарушения у беспозвоночных: эндокринология, тестирование и оценка. SETAC Press, Пенсакола, Флорида, стр. 23–106. Ле Кюрио-Бельфон, О., Фьеве, Б., Сералини, Г.Э., Матье, М., 2005. Кратковременная биоаккумуляция, циркуляция и метаболизм эстрадиола-17β в устрице Crassostrea gigas. J. Exp. Mar. Biol. Ecol. 325, 125–133. Le Curieux-Belfond, O., Moslemi, S., Mathieu, M., Séralini, GE, 2001. Метаболизм андрогенов у устрицы Crassostrea gigas: данные об активности 17β-HSD и характеристика ароматазоподобной активности, ингибируемой фармакологическими соединениями и Морской загрязнитель.J. Steroid Biochem. Мол. Биол. 78, 359–366. Ли, Л.Дж., Гарден, Р.В., Флойд, П.Д., Мороз, Т.П., Глисон, Дж. М., Свидлер, Дж. В., Паса-Толик, Л., Смит, Р. Д., 1999. Пептиды гормона откладки яиц в семействе Aplysiidae. J. Exp. Биол. 202, 2961–2973. Ли, К., Осада, М., Судзуки, Т., Мори, К., 1998. Изменения вителлина во время оогенеза и влияние эстрадиола на вителлогенез у тихоокеанской устрицы Crassostrea gigas. Инвертировать. Репрод. Dev. 33, 87–93. Любе П., Матье М., 1990. Эндокринные регуляции двустворчатых моллюсков.Аня. Биол. 29, 235–252. Матье, М., 1994. Эндокринный контроль углеводного обмена у моллюсков. Перспектива. Комп. Эндокринол. 471–474. Матье, М., Роббинс, И., Любет, П., 1991. Нейроэндокринология Mytilus edulis. Аквакультура. 94, 213–223. Мацумото, Т., Накамура, А.М., Мори, К., Акияма, И., Хиросе, Х., Такахаши, Ю., 2007. Рецептор эстрогена устрицы: клонирование кДНК и иммунолокализация. Gen. Comp. Эндокринол. 151, 195–201. Мацумото, Т., Осада, М., Осава, Ю., Мори, К., 1997. Профиль гонадного эстрогена и иммуногистохимическая локализация стероидогенных ферментов в устрицах и гребешках во время полового созревания.Комп. Биохим. Physiol. B 118, 811–817. Маттиссен, П., Килбрайд, Р., Мейсон, К., Пендл, М., Рис, Х., Уолдок, Р., 1999. Мониторинг восстановления бентического сообщества в реке Крауч после заражения ТБО. Отчет перед Министерством окружающей среды, транспорта и регионов Великобритании. Центр науки об окружающей среде, рыболовстве и аквакультуре, Бернхэм-он-Крауч, Великобритания, стр. 51. EPG 1/5/92. Маквей, М.Дж., Кук, Г.М., 2003. Ингибирование активности микросомальной 3β-гидроксистероиддегидрогеназы семенников крысы трибутилоловом.J. Steroid Biochem. Мол. Биол. 86, 99–105. Minier, C., Abarnou, A., Le Guellec, AM, Jaouen-Madoulet, A., Tutundjian, R., Bocquené, G., Leboulenger, F., 2006. Пилотное исследование мониторинга загрязнения, включающее химический анализ и измерения биомаркеров в устье Сены с использованием мидий зебры (Dreissena polymorpha). Environ. Toxicol. Chem. 25, 112–119. Минье, К., Кальто, Г., Лебуленжер, Ф., Хилл, Э.М., 2000. Исследование распространенности интерсекс-рыб в регионах Приморской Сены и Сассекса. Analusis 28, 801–806.Морсилло, Ю., Алабалат, А., Порте, С., 1999. Мидии как стражи оловоорганического загрязнения: биоаккумуляция и влияние на Р450-опосредованную активность ароматазы. Environ. Toxicol. Chem. 18, 1203–1208. Морсилло, Ю., Порте, К., 2000. Доказательства нарушения эндокринной системы у моллюсков (Ruditapes decussata), пересаженных в загрязненную трибутилоловом среду. Environ. Загрязнение. 107, 47–52.
I. Ketata et al. / Сравнительная биохимия и физиология, часть C 147 (2008) 261–270 Morcillo, Y., Ronis, M.J.J., Porte, C., 1998. Влияние трибутилолова на метаболизм тестостерона I фазы и титры стероидов моллюска (Ruditapes decussata). Акват. Toxicol. 42, 1–13. Мори, К., 1969. Действие стероида на устрицу-IV. Ускорение полового созревания самок Crassostrea gigas за счет эстрадиола-17β. Бык. Яп. Soc. Sci.Fish. 35, 1077–1079. Морс, Д.Е., 1984. Биохимическая и генная инженерия для улучшения производства морских ушек и других ценных моллюсков. Аквакультура 39, 263–282. Му Ю.М., Янасэ Т., Ниши Ю., Такаянаги Р., Goto, K., Nawata, H., 2001. Комбинированное лечение специфическими лигандами для системы ядерных рецепторов PPARγ: RXR заметно ингибирует экспрессию цитохрома P450arom в раковых клетках гранулезы человека. Мол. Клетка. Эндокринол. 181, 239–248. Нэгл, Г.Т., Пейнтер, С.Д., Бланкеншип, Дж. Э., 1989. Посттрансляционный процессинг в модельных нейроэндокринных системах: предшественники и продукты, которые координируют репродуктивную активность у Aplysia и Lymnaea, J. Neurosci. Res. 23, 359–370. Ницца, Е.П., Морритт, Д., Крейн, М., Thorndyke, M., 2003. Долгосрочные и трансгенерационные эффекты воздействия нонилфенола на ключевом этапе развития Crassostrea gigas. Возможные эндокринные нарушения? Mar. Ecol. Прогресс. Сер. 256, 256–300. Nishikawa, J.-I., Mamiya, S., Kanayama, T., Nishikawa, T., Shiraishi, F., Horiguchi, T., 2004. Вовлечение ретиноидного рецептора X в развитие импосекса, вызванного оловоорганическими соединениями брюхоногие моллюски. Environ. Sci. Technol. 38, 6271–6276. Обердёрстер, Э., Макклеллан-Грин, П., 2000. Нейропептид APGWamide вызывает импосекс у грязевых улиток, Ilyanassa устарел.Пептиды 21, 1323–1330. Обердёрстер, Э., Макклеллан-Грин, П., 2002. Механизмы индукции импосекса у грязевых улиток, Ильянасса устарело: TBT как ингибитор нейротоксина и ароматазы. Mar. Environ. Res. 54, 715–718. Oehlmann, J., Bauer, B., Minchin, D., Schulte-Oehlmann, U., Fioroni, P., Markert, B., 1998. Imposex у Nucella lapillus и интерсекс у Littorina littorea: межвидовое сравнение двух TBT- вызывает эффекты и их географическое единообразие. Hydrobiologia 378, 199–213. Oehlmann, J., Schulte-Oehlmann, U., Тиллманн, М., Маркерт, Б., 2000. Действие эндокринных разрушителей на переджаберных улиток (Mollusca: Gastropoda) в лаборатории. Часть I: бисфенол А и октилфенол как ксеноэстрогены. Экотоксикология 9, 383–397. Oetken, M., Bachmann, J., Schulte-Oehlmann, U., Oehlmann, J., 2004. Доказательства эндокринных нарушений у беспозвоночных. Int. Rev. Cytol. 236, 1–44. Оно, К.М., Осада, М., Мацутани, Т., Мори, К., Намура, Т., 1982. Профиль гонадного простагландина F2a во время полового созревания у устрицы Crassostrea gigas.Mar. Biol. Lett. 3, 223–230. Орн, С., Холбек, Х., Мадсен, Т.Х., Норргрен, Л., Петерсен, Г.И., 2003. Развитие гонад и выработка вителлогенина у рыбок данио (Danio rerio), подвергшихся воздействию этинилэстрадиола и метилтестостерона. Акват. Toxicol. 65, 397–411. Ортис-Заррагойтиа, М., Кахаравиль, М.П., 2005. Биомаркеры воздействия и эффектов, связанных с воспроизводством мидий, подвергшихся воздействию эндокринных разрушителей. Arch. Environ. Contam. Toxicol. 49, 1–9. Осада, М., Номура, Т., Мори, К., 1992. Влияние эстрогена и серотонина на высвобождение яиц из яичника гребешка in vitro.Nippon Suisan Gakkaishi 58, 223–227. Осада, М., Такамура, Т., Сато, Х., Мори, К., 2003. Синтез вителлогенина в яичнике гребешка Patinopecten yessoensis: контроль эстрадиолом-17β и центральной нервной системой. J. Exp. Zool. А 299, 172–179. Пазос, А.Дж., Матье, М., 1999. Влияние пяти природных гонадотропин-высвобождающих гормонов на клеточные суспензии гонад морских двустворчатых моллюсков: стимуляция синтеза гониальной ДНК. Gen. Comp. Эндокринол. 113, 112–120. Пек, М.Р., Лабади, П., Минье, К., Хилл, Э.М., 2007. Профилирование экологических и эндогенных эстрогенов у мидий данио (Dreissena polymorpha). Chemosphere 69, 1–8. Philips, D.J.H., 1978. Использование биологических индикаторных организмов для количественного определения хлорорганических загрязнителей в водной среде: обзор. Environ. Загрязнение. 16, 167–229. Пуинян, AM, Лабади, П., Хилл, Э.М., Осада, М., Кишида, М., Накаоб, Р., Новилло, А., Каллард, И.П., Ротчелл, Дж. М., 2006. Лабораторное воздействие 17β-эстрадиола не удалось индуцировать экспрессию гена рецептора вителлогенина и эстрогена
269
у морских беспозвоночных Mytilus edulis.Акват. Toxicol. 79, 376–383. Reis-Henriques, M.A., Le Guellec, D., Remy-Martin, J.P., Adessi, G.L., 1990. Исследования эндогенных стероидов морского моллюска Mytilus edulis, L. методом газовой хроматографии и масс-спектрометрии. Комп. Биохим. Physiol. В 95, 303–309. Рейс-Энрикес, М.А., Коимбра, Дж., 1990. Вариации уровней прогестерона в Mytilus edulis в течение годового репродуктивного цикла. Комп. Биохим. Physiol. А 95, 343–348. Riffeser, M., Hock, B., 2002. Уровни вителлогенина в гемолимфе мидий — подходящий биомаркер воздействия эстрогенов? Комп.Биохим. Physiol. С 132, 75–84. Ринчард, Дж., Дабровски, К., Гарсиа-Абиадо, М.А., Оттобре, Дж., 1999. Поглощение и истощение 17α-метилтестостерона в плазме во время индукции маскулинизации в мышечной ткани Esox masquinongy: влияние на стероиды в плазме и изменение пола. Стероиды 64, 518–525. Ронис, М.Дж., Мейсон, А.З., 1996. Метаболизм тестостерона барвинком (Littorina littorea) in vitro и in vivo: эффекты трибутилолова. Mar. Env. Res. 42, 161–166. Рубос, E.W., Vanheumen, W.R.A., 1994. Процессинг и высвобождение пептидов каудодорсальными клетками Lymnaea stagnalis во время цикла яйцекладки. Мозг. Res. 644, 83–89. Сантос, М.М., Филипе, Л., Кастро, К., Виейра, Миннесота, Микаэль, Дж., Морабито, Р., Массаниссо, П., Рейс-Энрикес, Массачусетс, 2005. Новое понимание механизма индукции импосекса у вертушка Nucella lapillus. Комп. Биохим. Physiol. С 141, 101–109. Schulte-Oehlmann, U., Bettin, C., Fioroni, P., Oehlmann, J., Stroben, E., 1995. Marisa cornuarietis (Gastropoda, Prosobranchia) Потенциальный биоиндикатор TBT для пресноводных сред.Экотоксикология 4, 372–384. Сегнер, Х., Кэролл, К., Фенске, М., Янссен, CR, Маак, Г., Паско, Д., Шеферс, К., Ванденберг, Г.Ф., Уоттс, М., Венцель, А., 2003. Выявление эндокринных разрушающих эффектов у водных позвоночных и беспозвоночных: отчет европейского проекта IDEA. Ecotoxicol. Environ. Безопасность 54, 302–314. Shi, H., Changjiang, H., Xiujuan, Y., Sixi, Z., 2005. Обновленная схема импосекса для (Cantharus cecillei) (Gastropoda: Buccinidae) и новый механизм, ведущий к стерилизации самок с импозексом.Mar. Biol. 146, 717–723. Сиа, А., Пеллерин, Дж., Амиард, Дж. К., Пеллетье, Э., Виглино, Л., 2003. Задержанный гаметогенез и уровни прогестерона у моллюсков с мягкой оболочкой (Mya arenaria) в связи с загрязнением in situ оловоорганическими соединениями и тяжелыми металлами. в реке Св. Лаврентия (Канада). Комп. Биохим. Physiol. С 135, 145–156. Смауи-Дамак, В., Ребай, Т., Бертет, Б., Хамза-Чаффай, А., 2006. Влияет ли загрязнение кадмием на размножение моллюска Ruditapes decussates? Годовое тематическое исследование. Комп. Биохим.Physiol. С 143, 252–261. Смит, А.Б., Спайкер, С., Ван Миннен, Дж., Берк, Дж. Ф., де Винтер, Ф., Ван Элк, Р., Гераертс, WPM, 1996. Экспрессия и характеристика моллюсков, родственного инсулину пептида VII из моллюска Lymnaea stagnalis. Неврология 70, 589–596. Смит, Б.С., 1971. Сексуальность у американской грязевой улитки (Nassarius obseletus) Сэй. Proc Malacol. Soc. Лондон. 39, 377–378. Смит С.А., Нейсон Дж., Кролл Р.П., 1997. Обнаружение иммунореактивности, подобной APGWamide, у морского гребешка Placopecten magellanicus.Нейропептиды 31, 155–165. Спунер, Н., Гиббс, П.Е., Брайан, Г.В., Гоуд, Л.Дж., 1991. Влияние трибутилолова на титры стероидов у самок волчонка Nucella lapillus и развитие импосекса. Mar. Env. Res. 32, 37–49. Стефано, Г.Б., Чжу, В., Мантионе, К., Джонс, Д., Саламон, Э., Чо, Дж. Дж., Кадет, П., 2003. 17β-эстрадиол подавляет ганглиозные микроглиальные клетки посредством высвобождения оксида азота: присутствие транскрипт рецептора эстрогена β. Нейроэндокринол. Lett. 24, 130–136. Тех, С.Дж., Кларк, С.Л., Браун, К.Л., Луома, С.